Activation of mast cells is characterized by dramatic changes in cell morphology, intracellular transport and exocytosis. All these processes depend on the activities of cytoskeletal filamentous structures – microtubules, microfilaments and intermediate filaments. Different regulatory pathways regulate cytoskeletal functions indispensable for mast cell degranulation. Microtubules of activated mast cells has been studied in various cellular models ranging from mouse bone marrow-derived mast cells (BMMCs) and isolated rat peritoneal mast cells (RPMCs) to mouse BMMC lines (e.g. MC/9), rat basophilic leukemia cell lines (e.g. RBL-2H3) or human must cell lines (e.g. HMC-1).
Structure of microtubules
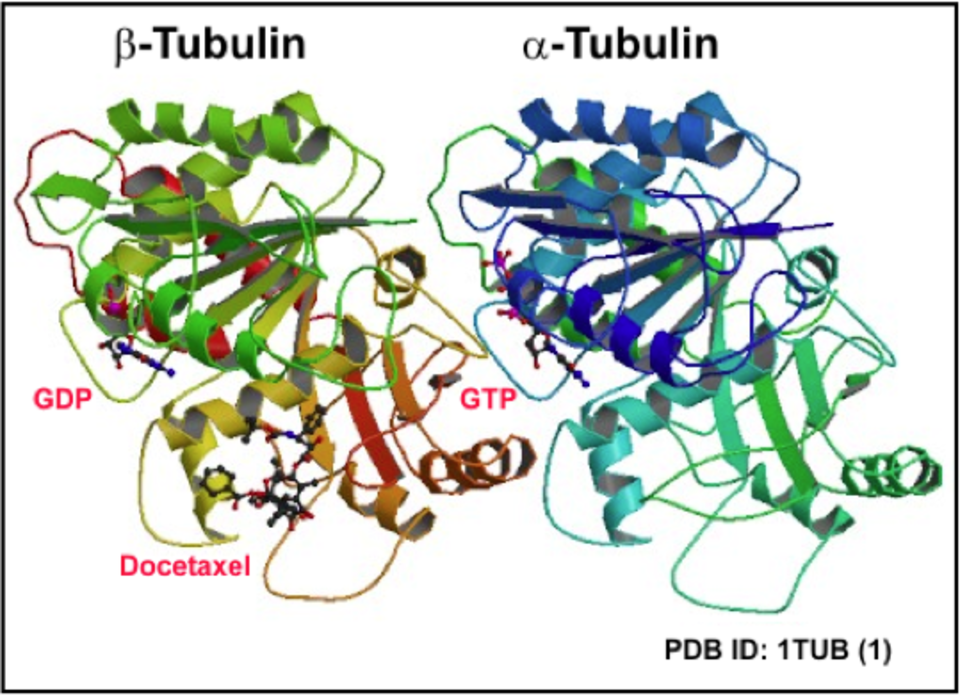
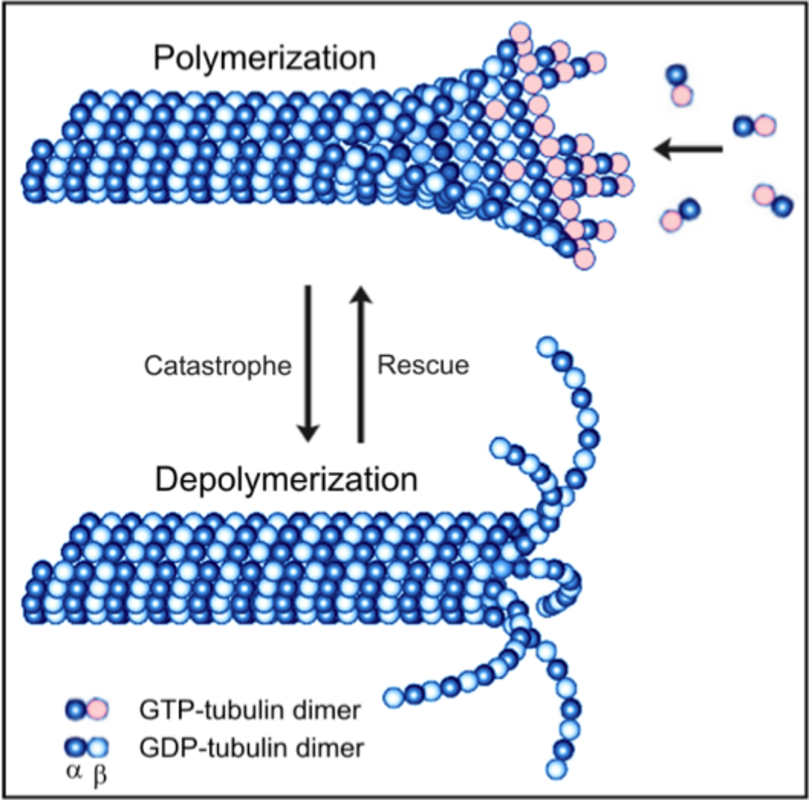
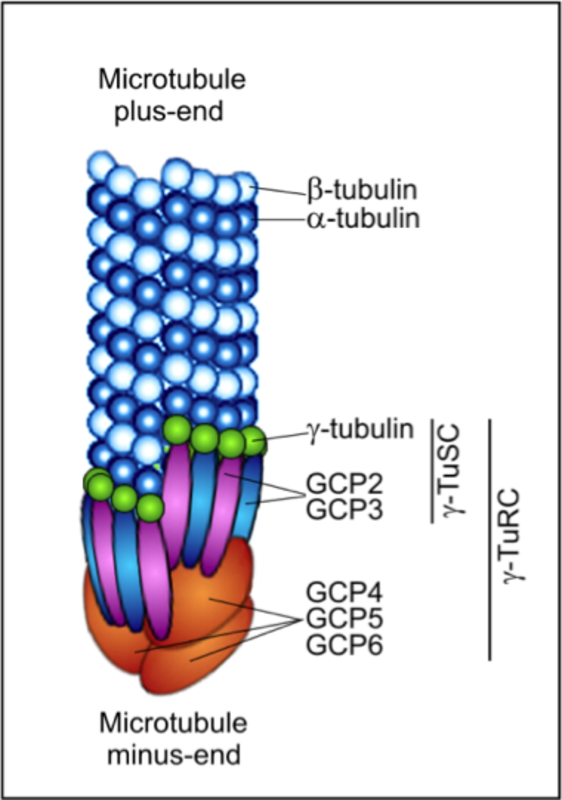
Microtubules are cytoskeletal polymers essential for the maintenance of cell shape, division, migration, compartmentalization and ordered intracellular transport powered by motor proteins. Microtubule is a cylinder with outer diameter 25 nm formed by 13 laterally bound protofilaments built by heterodimers of globular α- and β-tubulin subunits arranged in a head-to-tail manner (1). Microtubules are polar structures with two structurally distinct ends – a slow-growing minus end, exposing α-tubulin subunits, and a fast-growing plus end exposing β-tubulin subunits (2). Within the cells minus ends are stabilized by anchoring in microtubule-organizing centers (MTOCs), whereas plus ends are highly dynamic and switch between phases of growth and shrinkage. Each tubulin monomer binds one molecule of GTP, nonexchangeable in α-subunit (N-site) and exchangeable in β-subunit (E-site). Polymerization and depolymerization of microtubules are driven by hydrolysis of GTP on β-tubulin monomer. GTP exchange is necessary for the switch between phases of growth and shrinkage separated by catastrophe (growth to shrinkage transition) and rescue (from shortening to growth). This dynamic instability is an essential feature of microtubules that allows them to search through the cell for targets like cell cortex or actin cytoskeleton (3). The dynamics of microtubules can be regulated by incorporation of alternative tubulin isotypes – eight α-tubulin and seven β-tubulin phylogenetically conserved genes were identified in human (4), by post-translational modifications of tubulin subunits (mainly acetylation, tyrosination/detyrosination, polyglutamylation, polyglycylation and phosphorylation), and binding of microtubule associated proteins MAPs (5, 6). Microtubule dynamics can be also regulated by a variety of ligands, including microtubule drugs, i.e. docetaxel (7). The key regulator of microtubule nucleation is the γ-tubulin (8), a highly conserved minor member of the tubulin family concentrated in MTOCs. Within the cells γ-tubulin interacts with 6 GCPs (γ-tubulin complex proteins) to create small (γ-TuSC) and large (γ-TuRC) complexes that participate in microtubule nucleation (9). The plus-end tracking proteins (+TIPs), a diverse group of molecules that display dynamic accumulation at the ends of growing microtubules, represent important regulators of microtubule plus end dynamics (10).
Changes in microtubule organization in activated cells
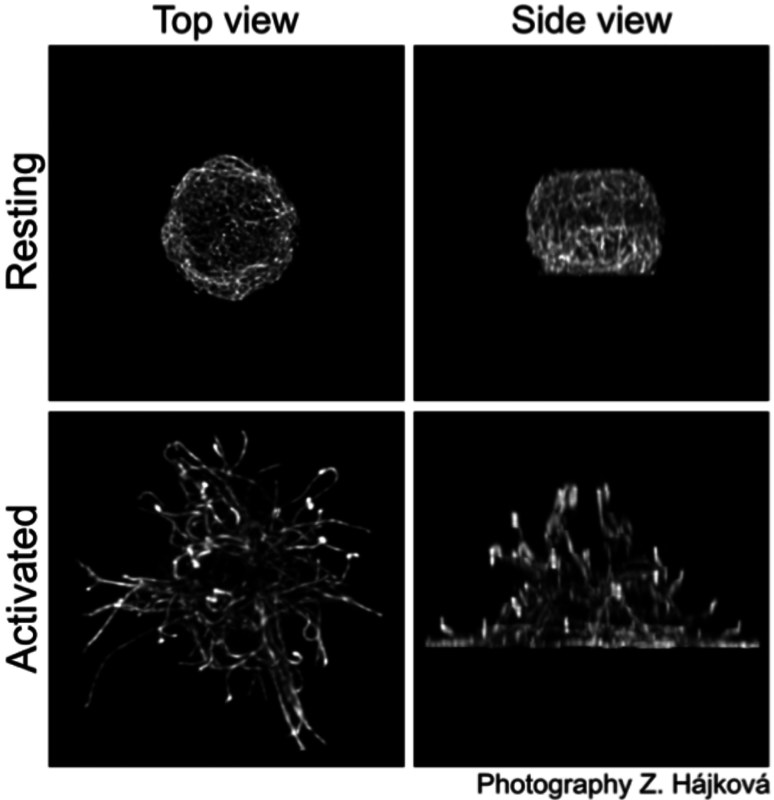
It is well established that microtubule drugs suppress exocytosis (11) and that movement of secretory granules in mast cells is dependent on intact microtubules (12-14). Activation of mast cells leads to changes in the topography of microtubules. FcεRI aggregation in BMMCs causes accumulation of microtubules in cell periphery (14, 15). Microtubules of BMMCs attached to fibronectin form protrusions in the course of mast cells activation; activation also modifies microtubule dynamics (16). The signaling pathway formed by tyrosine kinase Fyn, scaffolding adaptor protein Gab2 and Rho GTPase RhoA might play an important role in reorganization of microtubules after activation (14). It is well established that RhoA affects the dynamics of growing microtubules through its target mDia (17). Microtubule formation may also be influenced by +TIPs that interact at the cell periphery with cytoskeletal scaffold IQ domain GTPase-activating protein 1 (IQGAP1) which was found to sequester calmodulin and Rac GTPase (18). An important role in the regulation of microtubule formation and granule translocation in activated mast cells might also be played by RasGRPs that activate RhoA via PI3 kinase (19). Typical distribution of microtubules in resting and activated mouse BMMCs (16) is shown in the right panel.
Store-operated Ca2+ entry and microtubules
The store-operated Ca2+ entry (SOCE) is important for the replenishment of intracellular Ca2+ stores in endoplasmic reticulum (ER). One key component of SOCE is protein STIM1 (stromal interacting protein 1), the Ca2+-sensor associated with ER (20). After Ca2+ depletion from ER stores, STIM1 accumulates into ER subregions in close proximity to plasma membrane (10-25 nm) (21). STIM1 contains the SxIP sequence responsible for its direct interactions with end binding protein 1 (EB1; +TIP) and association with microtubules (20). Comet-like movement of STIM1 is observed by live-cell imaging in resting mast cells (16, 22). This movement is substantially reduced after mast cell activation (23, 24). Although microtubules could play a supportive role in SOCE signaling, several lines of evidence indicate that interaction of STIM1 with EB1 is not essential for the transport of STIM1 to plasma membrane (16).
γ-Tubulin in activated mast cells
Formation of microtubules is effectively regulated by microtubule nucleation. After mast cell activation, the Src-family kinases Lyn and Fyn form complexes containing γ-tubulin and proteins phosphorylated on tyrosine (15, 25). Src family kinases in mast cells form an essential part of signal transduction pathways located near the plasma membrane. However, they were also found in MTOC where they might regulate centrosomal recruitment of γ-tubulin (9). The Syk kinase, another important component of signaling pathways in activated mast cells, is capable of phosporylating α-tubulin (26) and interacts with γ-tubulin (15). Tyrosine kinases can thus play an important role in modulation of microtubules in activated mast cells (15, 27).
References:
| 1. | Nogales, E., S.G. Wolf, and K.H. Downing (1998) Structure of the alpha beta tubulin dimer by electron crystallography. Nature, 391: 199-203. |
| 2. | Nogales, E. and H.W. Wang (2006) Structural mechanisms underlying nucleotide-dependent self-assembly of tubulin and its relatives. Curr Opin Struct Biol, 16: 221-9. |
| 3. | Desai, A. and T.J. Mitchison (1997) Microtubule polymerization dynamics. Annu Rev Cell Dev Biol, 13: 83-117. |
| 4. | Luduena, R.F., Banerjee, A., (2008) The isotypes of tubulin: distribution and functional significance, in The Role of Microtubules in Cell Biology, Neurobiology and Oncology, Human Press: Totowa, NJ. 123-175. |
| 5. | Amos, L.A. and D. Schlieper (2005) Microtubules and maps. Adv Protein Chem, 71: 257-98. |
| 6. | Verhey, K.J. and J. Gaertig (2007) The tubulin code. Cell Cycle, 6: 2152-60. |
| 7. | Jordan, M.A. and K. Kamath (2007) How do microtubule-targeted drugs work? An overview. Curr Cancer Drug Targets, 7: 730-42. |
| 8. | Oakley, C.E. and B.R. Oakley (1989) Identification of gamma-tubulin, a new member of the tubulin superfamily encoded by mipA gene of Aspergillus nidulans. Nature, 338: 662-4. |
| 9. | Teixido-Travesa, N., J. Roig, and J. Luders (2012) The where, when and how of microtubule nucleation – one ring to rule them all. J Cell Sci |
| 10. | Akhmanova, A. and M.O. Steinmetz (2008) Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat Rev Mol Cell Biol, 9: 309-22. |
| 11. | Urata, C. and R.P. Siraganian (1985) Pharmacologic modulation of the IgE or Ca2+ ionophore A23187 mediated Ca2+ influx, phospholipase activation, and histamine release in rat basophilic leukemia cells. Int Arch Allergy Appl Immunol, 78: 92-100. |
| 12. | Martin-Verdeaux, S., et al. (2003) Evidence of a role for Munc18-2 and microtubules in mast cell granule exocytosis. J Cell Sci, 2003. 116: 325-34. |
| 13. | Smith, A.J., et al. (2003) Microtubule-dependent transport of secretory vesicles in RBL-2H3 cells. Traffic 4: 302-12. |
| 14. | Nishida, K., et al. (2005) FceRI-mediated mast cell degranulation requires calcium-independent microtubule-dependent translocation of granules to the plasma membrane. J Cell Biol, 170: 115-26. |
| 15. | Sulimenko, V., et al. (2006) Regulation of microtubule formation in activated mast cells by complexes of gamma-tubulin with Fyn and Syk kinases. J Immunol, 176: p. 7243-53. |
| 16. | Hájková, Z., et al. (2011) STIM1-directed reorganization of microtubules in activated mast cells. J Immunol, 186: 913-23. |
| 17. | Palazzo, A.F., et al. (2001) mDia mediates Rho-regulated formation and orientation of stable microtubules. Nat Cell Biol, 2001. 3: 723-9. |
| 18. | Logue, J.S., et al. (2011) AKAP220 protein organizes signaling elements that impact cell migration. J Biol Chem, 286: 39269-81. |
| 19. | Liu, Y., et al. (2007) An essential role for RasGRP1 in mast cell function and IgE-mediated allergic response. J Exp Med, 2007. 204: 93-103. |
| 20. | Roos, J., et al. (2005) STIM1, an essential and conserved component of store-operated Ca2+ channel function. J Cell Biol, 169: 435-45. |
| 21. | Cahalan, M.D. (2009) STIMulating store-operated Ca(2+) entry. Nat Cell Biol, 11: 669-77. |
| 22. | Grigoriev, I., et al. (2008) STIM1 is a MT-plus-end-tracking protein involved in remodeling of the ER. Curr Biol, 18: 177-82. |
| 23. | Baba, Y., et al. (2006), Coupling of STIM1 to store-operated Ca2+ entry through its constitutive and inducible movement in the endoplasmic reticulum. Proc Natl Acad Sci U S A, 2006. 103: 16704-9. |
| 24. | Calloway, N., et al. (2009) Molecular clustering of STIM1 with Orai1/CRACM1 at the plasma membrane depends dynamically on depletion of Ca2+ stores and on electrostatic interactions. Mol Biol Cell, 2009. 20: 389-99. |
| 25. | Dráberová, L., et al. (1999) Protein tyrosine kinase p53/p56(lyn) forms complexes with gamma-tubulin in rat basophilic leukemia cells. Int Immunol, 11: 1829-39. |
| 26. | Peters, J.D., et al. (1996) Syk, activated by cross-linking the B-cell antigen receptor, localizes to the cytosol where it interacts with and phosphorylates alpha-tubulin on tyrosine. J Biol Chem, 271: 4755-62. |
| 27. | Kukharskyy, V., et al. (2004) Complexes of gamma-tubulin with nonreceptor protein tyrosine kinases Src and Fyn in differentiating P19 embryonal carcinoma cells. Exp Cell Res, 298: 218-28. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}