Proper activity of cytoskeletal proteins is necessary for processes accompanying the activation of mast cells (e.g. changes in cell morphology, enhanced cell adhesion to various substrates, migration, and exocytosis). The cytoskeleton of mast cells is formed by three types of filamentous structures: microtubules consisting of αβ-tubulin dimers, microfilaments (F-actin) composed of actin, and intermediate filaments of vimentin-type. Microtubules and microfilaments are inherently polar structures with slow-growing minus ends and fast-growing plus ends. Research has mostly been focused on microfilaments and microtubules, while data on the involvement of intermediate filaments in mast cell activation are still very limited (1). The importance of cytoskeleton integrity for activity of mast cells is mostly based on evaluation of tests using cytoskeletal drugs.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Tubulin-targeting drugs
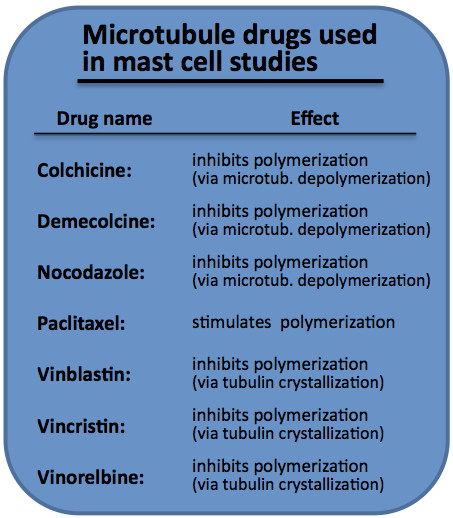
Tubulin-targeting drugs called also tubulin-binding agents (TBAs) are small-molecule inhibitors with diverse structures. They have been isolated from a broad range of species, including bacteria, marine sponges, molluscs and plants. At high concentrations, TBAs are broadly classified as microtubule-stabilizing or microtubule-destabilizing agents according to their effects on the mass of microtubule polymers (2). It is now apparent that TBAs can also exert their toxic effects in cancer cells at drug concentrations that disrupt microtubule dynamics without having a marked effect on microtubule polymer levels (3). TBAs are also commonly referred to as anti-mitotic agents because they cause mitotic arrest and cell death in tumour cells (3). Microtubule-stabilizing agents are particularly represented by taxanes and epothilones. Taxanes, e.g. paclitaxel, bind to β-tubulin, causing microtubule polymerization that blocks mitosis by virtue of kinetic stabilization of spindle microtubules (4). Epothilones are a new class of agents that, like paclitaxel, disrupt microtubule dynamics by binding to the β-tubulin subunit of αβ-tubulin and stabilize microtubules, inducing mitotic arrest and apoptosis. Epothilones and taxanes share overlapping but not identical binding sites on β-tubulin (5). Epothilone B and its derivatives (patupilone, ixabepilone and sagopilone ZK-EPO) are currently undergoing a stage of clinical development for the treatment of various tumours (6). Microtubule-destabilizing agents consist predominantly of drugs that act at the Vinca alkaloid and colchicine-binding sites (7). Vinca alkaloids (vincristine, vinblastine and vinorelbine) interact with the central portion of the β-tubulin subunit and thus prevent polymerization into microtubules (8). At high concentrations these drugs form so called tubulin paracrystals. Colchicine acts at a separate site on β-tubulin termed the colchicine-binding site where it depolymerises microtubules at high concentration and inhibits microtubule dynamics at low concentration (9). Apart from these natural products nocodazole has an effect similar to that of microtubule-destabilizing TBAs (10). The binding of nocodazole to tubulin is reversible and a rapid recovery follows upon its removal.
Actin-targeting drugs
The majority of the compounds targeting actin cytoskeleton and modulating its dynamics were isolated from terrestrial plants, fungi, bacteria, marine nudibranchs and marine sponges (11). These natural products are classified as F-actin-stabilizing or F-actin destabilizing drugs depending on their effect on actin cytoskeleton (11). Microfilament-stabilizing agents are primarily represented by phallotoxin and jasplakinolide/jaspamide families. The best known member of the phallotoxin family is phalloidin (12). It is generally coupled with fluorophores to stain actin filaments in immunofluorescence. Jasplakinolide also called jaspamide is an actin filament stabilizer that is also a potent inducer of actin polymerization (11, 13). Microfilament-destabilizing agents consist predominantly of the latrunculins and the cytochalasins. Latrunculins prevent the assembly of actin monomers through a highly specific sequestration of monomeric actin which therefore promotes depolymerization of microfilaments (13, 14). The binding of latrunculins is reversible. The cytochalasins have various effects on actin cytoskeleton. They inhibit association of actin subunits at plus ends by capping them. At high concentrations, cytochalasins also can sever actin filaments, sequester actin monomers or actin dimers (11). Swinholide A is another microfilament drug that specifically sequesters actin dimers and severs actin filaments (15).
Application of cytoskeletal drugs in mast cells studies
Microtubules and microfilaments influence a variety of processes during mast cells activation. Cytoskeletal drugs are thus used as useful research tools to investigate the role of cytoskeleton in particular signal transduction pathways. Microfilament-destabilizing drugs cytochalasin D and latrunculin helped to reveal the changes in total F-actin level in response to mast cell activation (16). Experiments with these drugs (16, 17) brought new findings on tyrosine phosphorylation levels and interactions between cross-linked IgE-FcεRI and Lyn kinase, and contributed in a decisive way to full understanding of the involvement of microfilaments in early mast cell activation processes. The use of latrunculin B or cytochalasins (B/C/D/E) and the microfilament stabilizer jasplakinolide provided data that greatly improved our knowledge on the role of F-actin in mast cell exocytosis (16, 18, 19). Microtubule-destabilizing drugs (nocodazole, colchicine, demecolcine, vinblastine, vinorelbine) and microtubule stabilizer taxol (paclitaxel) definitively showed that microtubules participate in mast cell degranulation (18-25). The importance of microtubules for the movement of secretory granules was proved in experiments using nocodazole and colchicine (18, 20). Cytoskeletal drugs (nocodazol, colchicine, latrunculin and jasplakinolide) confirmed the role of mast cell cytoskeleton in the store-operated Ca2+ entry (16, 21, 26-28). Cell migration depends on microfilament rearrangement. Pretreatment of cells with cytochalasin B leads to inhibition of mast cell chemotaxis (29, 30).
Usage of cytoskeletal drugs in mast cell experiments
| Drug | Colchicine | Cytochalasin B | Cytochalasin D | Demecolcine | Jasplakinolide | Latrunculin B | Nocodazole | Paclitaxel | Vinblastine | Vinorelbine |
| Concen- tration |
30-500 µM | 1-10 µM | 0.5-2 µM | 1-500 µM | 3-5 µM | 0.5-1 µM | 3-10 µM | 10-100 µM | 500 µM | 30-500 µg/ml |
| Time | 15-60 min | 15-30 min | 10-15 min | 30-60 min | 45 min-3 h | 15 min | 15-60 min | 30 min | 60 min | 10-20 min |
| Refer- ences |
(19-22, 30) | (19, 29, 30) | (16, 17) | (19, 22) | (18, 28) | (16, 18) | (21, 26, 27) | (18, 19) | (22) | (23, 24) |
References:
| 1. | Dráber, P., Sulimenko, V., and Dráberová, E. (2012). Cytoskeleton in mast cell signaling. Front Immunol. 3: 130. |
| 2. | Katsetos, C.D. and Dráber, P. (2012). Tubulins as therapeutic targets in cancer: from bench to bedside. Curr Pharm Des. 18: 2778-2792. |
| 3. | Kavallaris, M. (2010). Microtubules and resistance to tubulin-binding agents. Nat Rev Cancer. 10: 194-204. |
| 4. | Jordan, M.A., Toso, R.J., Thrower, D., and Wilson, L. (1993). Mechanism of mitotic block and inhibition of cell proliferation by taxol at low concentrations. Proc Natl Acad Sci U S A. 90: 9552-9556. |
| 5. | Nettles, J.H., Li, H., Cornett, B., Krahn, J.M., Snyder, J.P., and Downing, K.H. (2004). The binding mode of epothilone A on alpha,beta-tubulin by electron crystallography. Science. 305: 866-869. |
| 6. | Cortes, J. and Vidal, M. (2012). Beyond taxanes: the next generation of microtubule-targeting agents. Breast Cancer Res Treat. 133: 821-830. |
| 7. | Morris, P.G. and Fornier, M.N. (2008). Microtubule active agents: beyond the taxane frontier. Clin Cancer Res. 14: 7167-7172. |
| 8. | Rai, S.S. and Wolff, J. (1996). Localization of the vinblastine-binding site on beta-tubulin. J Biol Chem. 271: 14707-14711. |
| 9. | Jordan, M.A. and Wilson, L. (2004). Microtubules as a target for anticancer drugs. Nat Rev Cancer. 4: 253-265. |
| 10. | De Brabander, M.J., Van de Veire, R.M., Aerts, F.E., Borgers, M., and Janssen, P.A. (1976). The effects of methyl (5-(2-thienylcarbonyl)-1H-benzimidazol-2-yl) carbamate, (R 17934; NSC 238159), a new synthetic antitumoral drug interfering with microtubules, on mammalian cells cultured in vitro. Cancer Res. 36: 905-916. |
| 11. | Kustermans, G., Piette, J., and Legrand-Poels, S. (2008). Actin-targeting natural compounds as tools to study the role of actin cytoskeleton in signal transduction. Biochem Pharmacol. 76: 1310-1322. |
| 12. | Low, I. and Wieland, T. (1974). The interaction of phalloidin. Some of its derivatives, and of other cyclic peptides with muscle actin as studied by viscosimetry. FEBS Lett. 44: 340-343. |
| 13. | Spector, I., Braet, F., Shochet, N.R., and Bubb, M.R. (1999). New anti-actin drugs in the study of the organization and function of the actin cytoskeleton. Microsc Res Tech. 47: 18-37. |
| 14. | Coue, M., Brenner, S.L., Spector, I., and Korn, E.D. (1987). Inhibition of actin polymerization by latrunculin A. FEBS Lett. 213: 316-318. |
| 15. | Bubb, M.R., Spector, I., Bershadsky, A.D., and Korn, E.D. (1995). Swinholide A is a microfilament disrupting marine toxin that stabilizes actin dimers and severs actin filaments. J Biol Chem. 270: 3463-3466. |
| 16. | Frigeri, L. and Apgar, J.R. (1999). The role of actin microfilaments in the down-regulation of the degranulation response in RBL-2H3 mast cells. J Immunol. 162: 2243-2250. |
| 17. | Holowka, D., Sheets, E.D., and Baird, B. (2000). Interactions between Fc(epsilon)RI and lipid raft components are regulated by the actin cytoskeleton. J Cell Sci. 113: 1009-1019. |
| 18. | Nishida, K., Yamasaki, S., Ito, Y., Kabu, K., Hattori, K., Tezuka, T., Nishizumi, H., Kitamura, D., Goitsuka, R., Geha, R.S., Yamamoto, T., Yagi, T., and Hirano, T. (2005). Fc{epsilon}RI-mediated mast cell degranulation requires calcium-independent microtubule-dependent translocation of granules to the plasma membrane. J Cell Biol. 170: 115-126. |
| 19. | Urata, C. and Siraganian, R.P. (1985). Pharmacologic modulation of the IgE or Ca2+ ionophore A23187 mediated Ca2+ influx, phospholipase activation, and histamine release in rat basophilic leukemia cells. Int Arch Allergy Appl Immunol. 78: 92-100. |
| 20. | Smith, A.J., Pfeiffer, J.R., Zhang, J., Martinez, A.M., Griffiths, G.M., and Wilson, B.S. (2003). Microtubule-dependent transport of secretory vesicles in RBL-2H3 cells. Traffic. 4: 302-312. |
| 21. | Oka, T., Hori, M., and Ozaki, H. (2005). Microtubule disruption suppresses allergic response through the inhibition of calcium influx in the mast cell degranulation pathway. J Immunol. 174: 4584-4589. |
| 22. | Gillespie, E., Levine, R.J., and Malawista, S.E. (1968). Histamine release from rat peritoneal mast cells: inhibition by colchicine and potentiation by deuterium oxide. J Pharmacol Exp Ther. 164: 158-165. |
| 23. | Botana, L.M., Arnaez, E., Vieytes, M.R., Alfonso, A., Bujan, M.J., Louzao, M.C., and Cabado, A. (1992). Non-immunological release of histamine from rat mast cells elicited by antineoplastic agents. Cancer Chemother Pharmacol. 29: 495-498. |
| 24. | Estevez, M.D., Vieytes, M.R., Louzao, M.C., Alfonso, A., Vilarino, N., and Botana, L.M. (1997). The antineoplastic drug vinorelbine activates non-immunological histamine release from rat mast cells. Inflamm Res. 46: 119-124. |
| 25. | Nielsen, E.H. and Johansen, T. (1986). Effects of dimethylsulfoxide (DMSO), nocodazole, and taxol on mast cell histamine secretion. Acta Pharmacol Toxicol (Copenh). 59: 214-219. |
| 26. | Smyth, J.T., DeHaven, W.I., Bird, G.S., and Putney, J.W., Jr. (2007). Role of the microtubule cytoskeleton in the function of the store-operated Ca2+ channel activator STIM1. J Cell Sci. 120: 3762-3771. |
| 27. | Hájková, Z., Bugajev, V., Dráberová, E., Vinopal, S., Dráberová, L., Janáček, J., Dráber, P., and Dráber, P. (2011). STIM1-directed reorganization of microtubules in activated mast cells. J Immunol. 186: 913-923. |
| 28. | Oka, T., Sato, K., Hori, M., Ozaki, H., and Karaki, H. (2002). FcepsilonRI cross-linking-induced actin assembly mediates calcium signalling in RBL-2H3 mast cells. Br J Pharmacol. 136: 837-846. |
| 29. | Shimizu, T., Owsianik, G., Freichel, M., Flockerzi, V., Nilius, B., and Vennekens, R. (2009). TRPM4 regulates migration of mast cells in mice. Cell Calcium. 45: 226-232. |
| 30. | Kuehn, H.S., Radinger, M., Brown, J.M., Ali, K., Vanhaesebroeck, B., Beaven, M.A., Metcalfe, D.D., and Gilfillan, A.M. (2010). Btk-dependent Rac activation and actin rearrangement following FcepsilonRI aggregation promotes enhanced chemotactic responses of mast cells. J Cell Sci. 123: 2576-2585. |