Leishmania is an important cutaneous pathogen and more than 12 million people worldwide are currently infected. Almost 60,000 individuals die of complications associated with leishmaniasis and 2 million new infections are reported every year. In humans, different disease forms develop depending on the parasite strain. Visceral leishmaniasis is caused by L. donovani, whereas mucocutaneous disease results from inoculation of L. braziliensis. Cutaneous disease is caused by several strains, e.g. L. major, L. mexicana and L. tropica. In immunocompromised patients such as HIV-coinfected individuals, visceralization/fatal disease can also be observed upon infection with a dermatotropic strain. Although 350 million people are at risk of infection every day, an efficacious vaccine does not exist [von Stebut, 2007; Kronenberg et al., 2008]. Research in leishmaniasis will provide a better understanding of basic mechanisms for development of cellular immune responses and aims at the development of improved methods for therapy and prophylaxis.
{kind=link}
{kind=link}
{kind=link}
Leishmania life cycle:
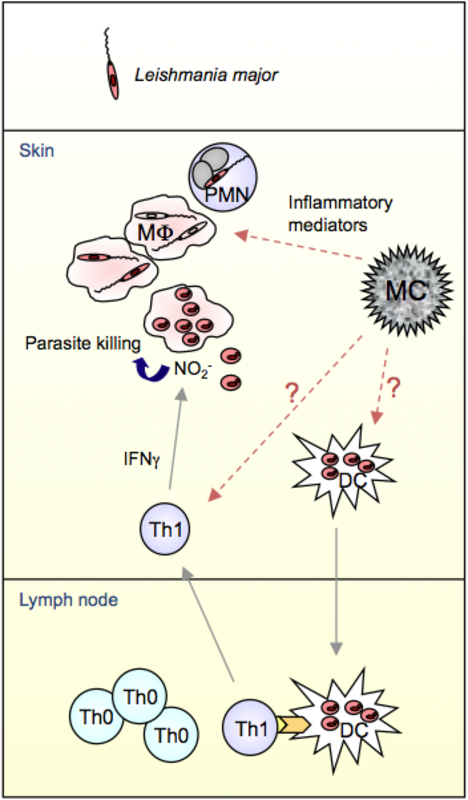
Leishmania parasites are inoculated into skin upon the bite of a sandfly. This leads to a rapid influx of neutrophils (PMN) to the wound site, which phagocytose L. major promastigotes. In the following, macrophages (MΦ) phagocytose infected PMN or they take up viable parasites released from apoptotic PMN. Inside the MΦ, promastigotes transform into the obligate intracellular amastigote life form. Free amastigotes are released from lysed MF and are able to infect dendritic cells (DC). Infected DC migrate to the draining lymph node (LN) and activate naïve T cells. Mast cells (MC) degranulate and release inflammatory mediators such as TNFα after infection and thereby recruit PMN to the site of infection [von Stebut et al., 2003]. L. major resistant strains like C57BL/6 develop a protective T helper (Th)1 response after infection. The life cycle is completed upon re-infection of sand flies after feeding at infected skin sites.
Protective immunity against L. major infection:
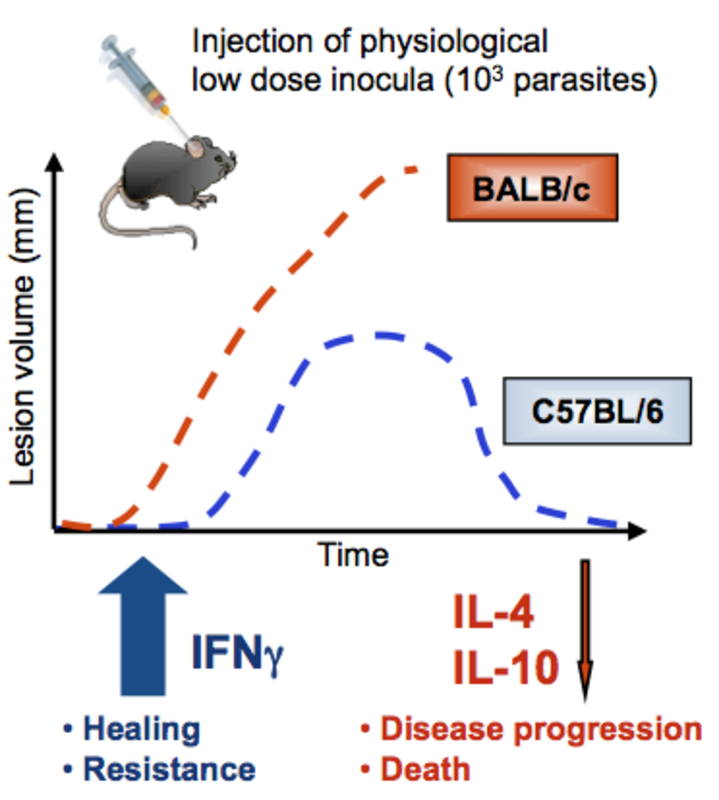
In leishmaniasis, protective, healing immune responses are largely dependent on effective IFNγ production mainly produced by Th1 CD4+ or cytotoxic T (Tc)1 CD8+ T cells [Sacks et al., 2002, Belkaid et al., 2002a]. This IFNγ mediates activation of infected MΦ to produce nitric oxide, which ultimately eliminates the parasites from the host. After development of the granulomatous lesion (‘oriental sore’), lesion involution, healing and life long immunity against re-infection is induced by these antigen-specific T cells.
Protective immunity against L. major is observed in most immune competent humans and the majority of mouse strains, including C57BL/6 mice. In contrast, immunocompromised patients (e.g. HIV-infected) and a few mouse strains (such as BALB/c mice) develop progressive disease due to Th2 responses and ultimately succumb to infection. Recently, we have shown that Th17 development is also associated with more progressive disease [Lopez Kostka et al., 2009], because increased levels of IL-17A induce persisting PMN infiltrates in progressive lesions [Tacchini-Cottier et al., 2000, Ribeiro-Gomes et al., 2004]. Depletion of PMN from L. major-infected BALB/c mice protected them from fatal disease [Tacchini-Cottier et al., 2000]. Finally, work from others has shown that regulatory T cells (Treg) promote parasite persistence via IL-10, which leads to worsening of disease outcome as well [Belkaid et al., 2002b], and we were able to demonstrate that epidermal dendritic Langerhans cells are responsible for Treg induction in this model [Kautz-Neu et al., 2011].
In search of the key factor/cell for the induction of early local inflammation against Leishmania, MC and MC- derived products are important candidates: First, MC are preferentially localized at the borders of the organism (skin, lung, gut) to contribute as sentinels of the immune system to local defence mechanisms. Second, MC produce a large array of proinflammatory mediators, some of which are stored in cytoplasmic granules and can be released within seconds after cell activation [Maurer et al., 1998]. And third, MC can be activated upon ligation of several pattern recognition receptors (e.g. TLR, complement receptors, CD48).
MC are responsible for survival in various models of acute bacterial and parasitic infections [Galli et al., 1999, Henz et al., 2001, Maurer et al., 2004, Mayerhofer et al., 2007, Marshall et al., 2004, Galli et al., 2008, Abraham et al., 2010]. Several authors have convincingly demonstrated that Leishmania-infected MC are activated, degranulated and released preformed TNFα both in vitro and in vivo [Bidri et al., 1997; Cangussu et al., 2009; Maurer et al., 2006]. Intradermal inoculation of physiologically relevant low doses of L. major resulted in local MC activation and MC-deficient KitwIKitw-v displayed significantly worsened disease with larger lesions, higher lesional parasite burdens, enhanced visceralization and predominant Th2 immune responses. Selective adoptive transfer of in vitro-derived MC to the skin of KitwIKitw-v mice repairing their MC deficiency resulted in the normalization of lesion development after infection. Finally, mechanistically, peritoneal cultured MC (PCMC) directly bound to immature DC and this cell-to-cell crosstalk resulted in an increased expression of the costimulators CD86, CD80 and CD40 on DC surfaces, indicating DC maturation [Dudeck et al., 2011]. The supernatants of MC/DC cocultures contained several T- cell modulating cytokines, e.g. IFNγ, IL-2, IL-6 and TGFβ. These results demonstrated that the crosstalk of DC with MC strongly impacts the subsequent capacity of DC to activate CD4+ T cells and to polarize the T cells towards a Th1 or Th17 response. This was also true for T cell responses against L. major.
Literature:
| Abraham SN, St John AL. Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol. 2010; 10:440-52 |
| Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature. 2002; 420:502-7b. |
| Belkaid Y, von Stebut E, Mendez S, Lira R, Caler E, Bertholet S, Udey MC, Sacks D. CD8+ T cells are required for primary immunity in C57BL/6 mice following low-dose, intradermal challenge with Leishmania major. J Immunol. 2002; 168:3992-4000a. |
| Bidri M, Vouldoukis I, Mossalayi MD, Debre P, Guillosson JJ, Mazier D, Arock M. Evidence for direct interaction between mast cells and Leishmania parasites. Parasite Immunol. 1997; 19:475-83. |
| Cangussú SD, Souza CC, Campos CF, Vieira LQ, Afonso LC, Arantes RM. Histopathology of Leishmania major infection: revisiting L. major histopathology in the ear dermis infection model. Mem Inst Oswaldo Cruz. 2009; 104:918-22. |
| Dudeck A, Suender CA, Kostka SL, von Stebut E, Maurer M. Mast cells promote Th1 and Th17 responses by modulating dendritic cell maturation and function. Eur J Immunol. 2011; 41:1883-93a; |
| Galli SJ, Grimbaldeston M, Tsai M. Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nat Rev Immunol. 2008; 8:478-86. |
| Galli SJ, Mauer M, Lantz CS. Mast cells as sentinels of innate immunology. Curr Opin Immunol. 1999; 11:53-59. |
| Henz B, Maurer M, Lippert U, Worm M, Babina M. Mast cells as initiators of immunity and host defense. Exp Dermatol. 2001; 10:1-10. |
| Kautz-Neu K, Noordegraaf M, Dinges S, Bennett CL, Clausen BE, von Stebut E: Langerhans cells are important negative regulators of the anti-Leishmania response. J Exp Med. 2011; 208:885-91; |
| Kronenberg K, Brosch S, von Stebut E. Vaccinations against cutaneous Leishmania infection. G Ital Dermatol Venereol. 2008; 143:125-37. |
| Lopez Kostka S, Dinges S, Griewank K, Iwakura Y, Udey MC, von Stebut E. IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J Immunol. 2009; 182:3039-46. |
| Marshall JS. Mast-cell responses to pathogens. Nat Rev Immunol. 2004; 4:787-99. |
| Maurer M, Echtenacher B, Hultner L, Kollias G, Mannel DN, Langley KE, Galli SJ. The c-kit ligand, stem cell factor, can enhance innate immunity through effects on mast cells. J Exp Med. 1998; 188:2343-8. |
| Maurer M, Lopez Kostka S, Siebenhaar F, Moelle K, Metz M, Knop J, von Stebut E. Skin mast cells control T cell-dependent host defense in Leishmania major infections. FASEB J. 2006; 20: 2460-7. |
| Maurer M, von Stebut E. Macrophage inflammatory protein-1. Int J Biochem Cell Biol. 2004; 36:1882-6. |
| Mayerhofer M, Aichberger KJ, Florian S, Valent P. Recognition sites for microbes and components of the immune system on human mast cells: relationship to CD antigens and implications for host defense. Int J Immunopathol Pharmacol. 2007; 20:421-34. |
| Ribeiro-Gomes FL, Otero AC, Gomes NA, Moniz-De-Souza MC, Cysne-Finkelstein L, Arnholdt AC, Calich VL, Coutinho SG, Lopes MF, DosReis GA. Macrophage interactions with neutrophils regulate Leishmania major infection. J Immunol. 2004; 172:4454-62. |
| Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002; 2:845-58. |
| Tacchini-Cottier F, Zweifel C, Belkaid Y, Mukankundiye C, Vasei M, Launois P, Milon G, Louis JA. An immunomodulatory function for neutrophils during the induction of a CD4+ Th2 response in BALB/c mice infected with Leishmania major. J Immunol. 2000; 165:2628-36. |
| Von Stebut E, Metz M, Milon G, Knop J, Maurer M. Early macrophage influx to sites of cutaneous granuloma formation is dependent on MIP-1α/β released from neutrophils recruited by mast cell-derived TNF-α. Blood 2003; 101:210-5. |
| Von Stebut E. Immunology of cutaneous leishmaniasis: the role of mast cells, phagocytes and dendritic cells for protective immunity. Eur J Dermatol. 2007; 17:115-22. |