Transmembrane adaptor proteins (TRAPs) facilitate activation within signal transduction pathways but lack both intrinsic enzymatic and transcriptional activities. They are characterized by a short extracellular domain, a single transmembrane domain and a cytoplasmic tail with various motifs serving as anchors for cytoplasmic signaling molecules. TRAPs also often contain a juxtamembrane palmitoylation motif, CXXC, that determines their solubility in nonionic detergents, distribution in the plasma membrane and functional properties (Table 1). Five of the TRAPs, namely LAT (Linker for Activation of T cells), NTAL (Non-T-cell Activation Linker), PAG [Phosphoprotein Associated with Glycosphingolipid-enriched membrane microdomains (GEMs)], LAX (Linker for Activation of X cells, X stands for “to be defined”) and GAPT (Grb2-binding Adaptor, Transmembrane), have been identified in mast cells and four of them (LAT, NTAL, LAX and PAG) have been shown to be involved in FcεRI and c-Kit signaling. Several reviews (1-7) deal with properties of TRAPs as well as their functions.
{kind=link}
{kind=link}
LAT
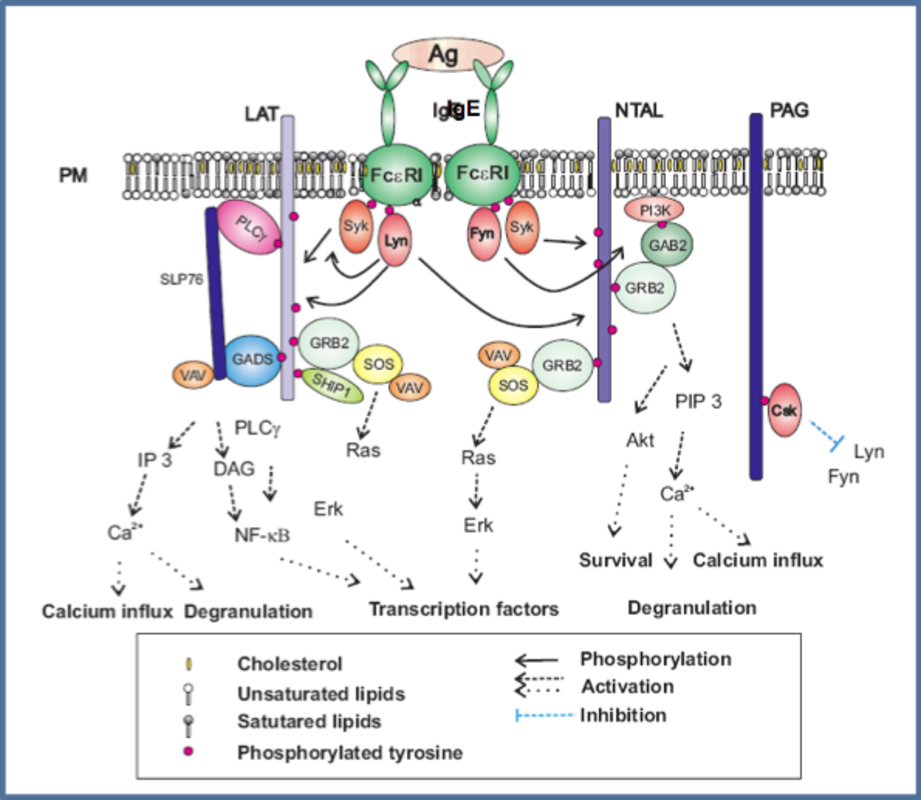
One of the most extensively studied TRAPs is LAT that was initially characterized as a 36-38 kDa protein prominently tyrosine-phosphorylated upon T-cell activation (8). Later it was also described in other cells of the immune system, including natural killer (NK) cells, platelets, megakaryocytes, immature B cells and mast cells. Like other TRAPs, LAT is composed of a short extracellular domain, a transmembrane domain which is followed by palmitoylation sites (9) and a cytoplasmic domain that possesses 9 tyrosine residues, which are conserved in humans, mice and rats; five of these tyrosines can be phosphorylated and serve as binding sites for SH2 domain-containing proteins (Table 1). Upon FcεRI activation, LAT is rapidly phosphorylated (10) and forms complexes with intracellular adaptor proteins such as SLP-76, Grb2, Gads, phospholipase Cγ1 and the guanine nucleotide exchange factor (GEF), Vav1 (11-14). LAT plays a key role in FcεRI-induced mast cell activation. Mast cells derived from bone marrow of Lat -/- mice exhibited severe decrease in the phosphorylation of numerous proteins, degranulation, and cytokine production. Lat -/- mice have normal numbers of mature mast cells but are resistant to IgE-mediated passive systemic anaphylaxis (10).
NTAL/LAB/LAT2
NTAL also termed LAT2 (for its domain similarity with LAT) or linker for activation of B cells (LAB) was identified as a 30 kD phosphorylated protein in myeloid cell lines (15) and simultaneously by search in human genome database for molecules with Grb2-binding motifs (YXN, where X stands for any amino acid), putative transmembrane domain with a stretch of hydrophobic residues and potential palmitoylation sites (16). In activated mast cells, NTAL is rapidly phosphorylated by Syk, Lyn and/or c-kit, where each kinase phosphorylates different tyrosines (17). In comparison to wild-type mast cells, Ntal -/- mast cells show enhanced degranulation, calcium mobilization, PLCγ and Erk phosphorylation as well as cytokine production. In vivo studies showed that mice with NTAL knock-out exhibited significantly enhanced passive systematic anaphylaxis. All these findings suggest that NTAL, unlike LAT, has a negative regulatory role in mast cells signaling (18,19). On the other hand, mice lacking both LAT and NTAL exhibited deeper inhibition of FcεRI signaling pathways than Lat -/- cells suggesting than NTAL plays also a positive regulatory role in mast cell activation.
PAG/CBP
PAG also termed Csk-binding protein (Cbp) was first identified in T-cells (20) and neonatal rat brain (21). Similarly to LAT and NTAL, PAG is palmitoylated and localized in detergent resistant membranes, but in contrast to other TRAPs none of its tyrosines is involved in binding of Grb2 adaptor. On the other hand, PAG possesses two proline rich sequences that could serve as binding sites for SH3 domain-containing proteins (20). Whereas PAG is constitutively phosphorylated in primary T cells and its phosphorylation decreases after T-cell receptor crosslinking (20) in mast cells PAG exhibits increased phosphorylation after FcεRI triggering (22,23). PAG in mast cells is phosphorylated by Lyn; in its absence PAG phosphorylation decreases and this leads to lower binding of Csk (22). Overexpression of PAG causes reduced phosphorylation of FcεRI β and γ subunits, degranulation and calcium release; these data suggest that PAG serves as a negative regulator of mast cell activation (20).
LAX
This adaptor has been discovered by searching human genome database for tyrosine motifs found in LAT (24). LAX is phosphorylated in mast cells in response to FcεRI activation and interacts with Grb2 and p85. LAX-deficient mast cells are hyperresponsive after FcεRI triggering. They exhibit enhanced degranulation, cytokine production, survival and activation of PI3K, Akt and p38 MAPK. The observed hyperresponsiveness seems to be a consequence of reduced expression of NTAL, observed after sensitization of mast cells with anti-dinitrophenyl IgE (25).
GATP
Its role in mast cell signaling is still enigmatic. It was discovered by in silico screening for novel adaptor proteins in human genome database. GAPT is expressed in mast cells, dendritic cells and murine B220+ spleen cells, but is missing in macrophages and T-cells (26). In contrast to other TRAPs described above, GAPT is not phosphorylated upon FcεRI-mediated activation, even though it possesses seven tyrosine residues, four of which with Grb2-binding motif.
References:
| 1. | Alvarez-Errico D, Lessmann E, & Rivera J (2009) Immunol. Rev., 232, 195-217. |
| 2. | Draber P, Halova I, Levi-Schaffer F, & Draberova L (2011) Front Immunol., 2, 95. |
| 3. | Fuller DM & Zhang W (2009) Immunol. Rev., 232, 72-83. |
| 4. | Fuller DM, Zhu M, Ou-Yang CW, Sullivan SA, & Zhang W (2011) Immunol. Res., 49, 97-108. |
| 5. | Hořejší V, Zhang W, & Schraven B (2004) Nat. Rev. Immunol., 4, 603-616. |
| 6. | Rivera J (2005) Trends Immunol., 26, 119-122. |
| 7. | Simeoni L, Kliche S, Lindquist J, & Schraven B (2004) Curr. Opin. Immunol., 16, 304-313. |
| 8. | June CH, Fletcher MC, Ledbetter JA, & Samelson LE (1990) J. Immunol., 144, 1591-1599. |
| 9. | Zhang W, Trible RP, & Samelson LE (1998) Immunity., 9, 239-246. |
| 10. | Saitoh S, Arudchandran R, Manetz TS, Zhang W, Sommers CL, Love PE, Rivera J, & Samelson LE (2000) Immunity., 12, 525-535. |
| 11. | Arudchandran R, Brown MJ, Peirce MJ, Song JS, Zhang J, Siraganian RP, Blank U, & Rivera J (2000) J. Exp. Med., 191, 47-60. |
| 12. | Finco TS, Kadlecek T, Zhang W, Samelson LE, & Weiss A (1998) Immunity., 9, 617-626. |
| 13. | Ishiai M, Kurosaki M, Inabe K, Chan AC, Sugamura K, & Kurosaki T (2000) J. Exp. Med., 192, 847-856. |
| 14. | Zhang W, Trible RP, Zhu M, Liu SK, McGlade CJ, & Samelson LE (2000) J. Biol. Chem., 275, 23355-23361. |
| 15. | Brdička T, Imrich M, Angelisová P, Brdičková N, Horváth O, Spička J, Hilgert I, Lusková P, Dráber P, Novák P, Engels N, Wienands J, Simeoni L, Österreicher J, Aguado E, Malissen M, Schraven B, & Hořejsi V (2002) J. Exp. Med., 196, 1617-1626. |
| 16. | Janssen E, Zhu M, Zhang W, Koonpaew S, & Zhang W (2003) Nat. Immunol., 4, 117-123. |
| 17. | Iwaki S, Jensen BM, & Gilfillan AM (2007) Int. J. Biochem. Cell Biol., 39, 868-873. |
| 18. | Volná P, Lebduška P, Dráberová L, Šimová S, Heneberg P, Boubelík M, Bugajev V, Malissen B, Wilson BS, Hořejši V, Malissen M, & Dráber P (2004) J. Exp. Med., 200, 1001-1013. |
| 19. | Zhu M, Liu Y, Koonpaew S, Granillo O, & Zhang W (2004) J. Exp. Med., 200, 991-1000. |
| 20. | Brdička T, Pavlištová D, Leo A, Bruyns E, Kořínek V, Angelisová P, Scherer J, Shevchenko A, Hilgert I, Černý J, Drbal K, Kuramitsu Y, Kornacker B, Hořejší V, & Schraven B (2000) J. Exp. Med., 191, 1591-1604. |
| 21. | Kawabuchi M, Satomi Y, Takao T, Shimonishi Y, Nada S, Nagai K, Tarakhovsky A, & Okada M (2000) Nature, 404, 999-1003. |
| 22. | Odom S, Gomez G, Kovarova M, Furumoto Y, Ryan JJ, Wright HV, Gonzalez-Espinosa C, Hibbs ML, Harder KW, & Rivera J (2004) J. Exp. Med., 199, 1491-1502. |
| 23. | Ohtake H, Ichikawa N, Okada M, & Yamashita T (2002) J. Immunol., 168, 2087-2090. |
| 24. | Zhu M, Janssen E, Leung K, & Zhang W (2002) J. Biol. Chem., 277, 46151-46158. |
| 25. | Zhu M, Rhee I, Liu Y, & Zhang W (2006) J. Biol. Chem., 281, 18408-18413. |
| 26. | Liu Y & Zhang W (2008J. Leukoc. Biol., 84, 842-851. |