General Description and Physiological Function
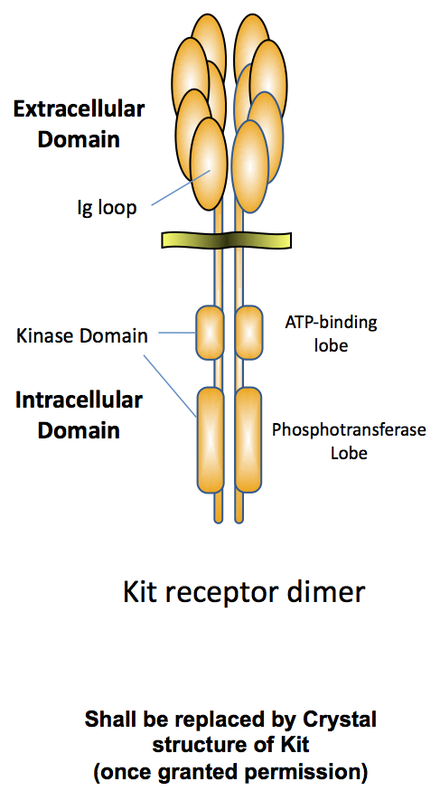
KIT is a transmembrane receptor, member of the receptor tyrosine kinase family and structurally related to the PDGF, CSF-1 (M-CSF) and FLT3 receptors. It is encoded by the c-kit gene, the cellular homolog of v-kit Hardy-Zuckerman 4 feline sarcoma viral oncogene. KIT and the PDGFr family members share a common structure made of an extracellular domain that contains five Ig loops, a single transmembrane region, an intracellular domain that contains the catalytic domain split in two by a kinase insert peptide. In the mouse the c-kit gene corresponds to the W (White-spotting) locus. KIT is the receptor for Stem Cell Factor (SCF). It has a restricted pattern of expression. It is expressed and is required in hematopoietic cells, melanocytes, germ cells and interstitial cells of Cajal. In addition KIT is expressed in populations of neurons in the brain, in the lung and is also a marker for several stem-cell populations.
Most of the knowledge of KIT’s physiological function comes from the study of W mutant mice (Lev 1994). These mice are either c-kit complete loss-of-fonction mutants or partial mutants with reduced expression or reduced catalytic activity (hypomorphs). Mice devoid of c-kit die around birth from macrocytic anemia. They do not have melanocytes in the skin, they are devoid of germ cells in the gonads, and lack mast cells in tissues and interstitial cells of Cajal in the digestive track. These defects are directly attributed to unique fonctions of KIT in the early development of cell lineages of diverse embryonic origin. KIT is required for primordial germ cells migration and proliferation, for the migration and expansion of melanoblasts, for normal hematopoieisis especially for the generation of red blood cells and mast cells. KIT is also required later on following early differentiation for melanocytes and mast cells function in the skin.
{kind=link}
KIT and Mast cells
In the hematopoietic system, KIT receptor is expressed in most early progenitors including in the stem-cell compartment (Broudy 1997). In most lineages, expression of KIT is lost during differentiation. Mast cells are an exception as they maintain KIT expression throughout all differentiation and maturation steps. Mouse mast cells can be generated in the presence of IL-3 or SCF (KIT-ligand). Human mast cells can only be generated in vitro upon continuous stimulation of SCF (Arock, 2008).
SCF and its receptor KIT have pleiotropic functions in mast cells, regulating cell survival, differenciation, proliferation, and modulating the activation of mast cells through Fc epsilon RI receptors (Galli 1995).
Signaling downstream of KIT
Binding of SCF to KIT receptor induces dimerization and activation of KIT. Several Tyr residues located (for human receptor) in the juxtamembrane (Y568 and Y570), in the kinase insert region (Y702, Y721, Y730) and in the C-terminus tail (Y736) become docking sites that recruit many signaling proteins to the receptor complex. Major signaling pathways activated downstream of KIT include the Ras/Erk1/2, the PI3-Kinase/AKT, PLCg and PKC pathways (Ronnstrand 2004). Other effectors include Src-family kinases, Fes family kinases, Stat1, Stat3, Stat5, Vav, Rac, p38 and Jnk.
Kit receptor signaling is maintained under control by multiple regulatory processes. Among these, the tyrosine phosphatase Shp1 (Paulson 1996), the adaptors Lnk (Takaki 2002), SOCS1 and 6 proteins (de Sepulveda 1999 and Bayle 2004, respectively) and Cbl (Zeng 2005) play critical roles.
In the mouse model, deficiency of the PI3-kinase regulatory sub-unit p85 (Krishnan 2012), of the adaptor Gab2 (Nishida 2002), of the tyrosine phosphatase Shp2 (Sharma 2012) or of the exchange-factor Swap70 (Sivalenka 2004) results in impaired KIT signaling and mast cell development and/or function.
Kit mutations and Mastocytosis
Mutations of c-kit gene are responsible for human pathologies. Heterezygous loss-of-function mutations result in Piebaldism (Spritz 1992), a pigmentation disease due to a melanocyte migration defect (similar to the coat color defects of W/+ mice). On the other hand, gain-of-function mutations occur in neoplastic disease such as gastro-intestinal stromal tumors, acute myeloid leukemia, germ-cell tumors, melanoma and mastocytosis (Ashman 2012). Kit mutations cluster in three hot-spots in the protein: in the catalytic domain, in the regulatory intracellular juxtamembrane domain and in the extracellular fifth Ig loop.
Mastocytosis is a rare multisymptomatic disorders characterized by the abnormal growth and accumulation of hyper-activated mast cells in one or more organs (Pardanani 2012). The disease is heterogeneous with child, adults and rare familial forms, and ranging from symptoms limited to the skin to mast cell leukemia. About 85% of patients have a somatic mutation of c-kit (Sperr 2012). In adults, the vast majority of the mutations are a point mutation in the catalytic kinase domain of KIT at position 816 replacing a conserved Asp residue. In young children, half of the patients have the same mutation while the other half have mutations in the extracellular domain of KIT receptor (Bodemer 2010; Yang 2010).
Pharmacology
KIT receptor is part of the protein kinase family and is recognized as a therapeutic target in GIST and mastocytosis. ATP-mimetics inhibitors have been selected to target KIT receptor. The first such molecule identified is imatinib, also called Gleevec/Glivec, a BCR-ABL, PDGFR and KIT inhibitor (Zermatti 2003). Imatinib inhibits wild-type Kit receptor and several Kit mutants in the micromolar range. However, the most common KIT mutant found in mastocytosis shows resistance to Imatinib and to several other Kit inhibitors. Nilotinib, Masitinib, Dasatinib, Sunitinib, Axitinib are second generation KIT inhibitors. Several small pharmacological inhibitors that target KIT are now available in clinic.
Gene/Protein Links
| Symbol: | KIT |
| Symbol: | CD117 |
| Symbol: | SCFR |
| PDB: | www.rcsb.org/pdb/home/home.do 1PKG |
| Uniprot: | www.uniprot.org P10721 (human KIT) |
| Uniprot: | www.uniprot.org P05532 (mouse) |
| Pfam: | pfam.sanger.ac.uk/family/PF00047 Immunoglobulin |
| Pfam: | pfam.sanger.ac.uk/family/PF07714 Protein Tyrosine Kinase |
| Genecards: | www.genecards.org KIT |
| Ensembl: | www.ensembl.org ENSG00000157404 |
| Protein Atlas: | www.proteinatlas.org ENSG00000157404 |
| OMIM: | omim.org/entry/164920 |
Animal Models
| W mice (Many alleles): | www.ncbi.nlm.nih.gov.gate1.inist.fr/pubmed/8252632 B6.129S2(Cg)-Fcer1a<tm1Knt>/J |
| GIST model KITΔ558 KI: | www.ncbi.nlm.nih.gov/pubmed/12754375 |
| GIST model KIT K641E KI: | www.ncbi.nlm.nih.gov/pubmed/16061643 |
| Mastocytosis models: | Zapulla et al. J Exp Med 2005 www.ncbi.nlm.nih.gov/pubmed/16352739 Gerbaulet et al Blood 2011 |
References:
| Arock, M., et al., Ex vivo and in vitro primary mast cells. Methods Mol Biol, 2008. 415: p. 241-54. |
| Ashman, L.K. and R. Griffith, Therapeutic targeting of c-KIT in cancer. Expert Opin Investig Drugs, 2012. |
| Bayle J, Letard S, Dubreuil P and De Sepulveda P. (2004) Suppressor of cytokine signaling 6 associates with KIT and regulates KIT receptor signaling. J. Biol. Chem. 279, 12249-12259. |
| Bodemer, C., et al., Pediatric mastocytosis is a clonal disease associated with D816V and other activating c-KIT mutations. J Invest Dermatol, 2010. 130(3): p. 804-15. |
| Broudy, V.C., Stem cell factor and hematopoiesis. Blood, 1997. 90(4): p. 1345-64. |
| De Sepulveda P., Klaus Okkenhaug, Jose La Rose, Patrice Dubreuil and Robert Rottapel (1999) Socs1 binds to multiple signalling proteins and suppresses Steel factor-dependent proliferation. EMBO J. 18, 904-915. |
| Dubreuil, P., et al., Masitinib (AB1010), a potent and selective tyrosine kinase inhibitor targeting KIT. PLoS One, 2009. 4(9): p. e7258. |
| Galli, S.J., et al., Regulation of mouse and human mast cell development, survival and function by stem cell factor, the ligand for the c-kit receptor. Int Arch Allergy Immunol, 1995. 107(1-3): p. 51-3. |
| Krishnan, S., et al., p85beta regulatory subunit of class IA PI3 kinase negatively regulates mast cell growth, maturation, and leukemogenesis. Blood, 2012. 119(17): p. 3951-61. |
| Lev, S., et al., Steel factor and c-kit protooncogene: genetic lessons in signal transduction. Crit Rev Oncog, 1994. 5(2-3): p. 141-68. |
| Nishida, K., et al., Requirement of Gab2 for mast cell development and KitL/c-Kit signaling. Blood, 2002. 99(5): p. 1866-9. |
| Pardanani, A., Systemic mastocytosis in adults: 2012 Update on diagnosis, risk stratification, and management. Am J Hematol, 2012. 87(4): p. 401-11. |
| Paulson, R.F., et al., Signalling by the W/Kit receptor tyrosine kinase is negatively regulated in vivo by the protein tyrosine phosphatase Shp1. Nat Genet, 1996. 13(3): p. 309-15. |
| Ronnstrand, L., Signal transduction via the stem cell factor receptor/c-Kit. Cell Mol Life Sci, 2004. 61(19-20): p. 2535-48. |
| Sharma, N., et al., SH2 domain-containing phosphatase 2 is a critical regulator of connective tissue mast cell survival and homeostasis in mice. Mol Cell Biol, 2012. 32(14): p. 2653-63. |
| Sivalenka, R.R. and R. Jessberger, SWAP-70 regulates c-kit-induced mast cell activation, cell-cell adhesion, and migration. Mol Cell Biol, 2004. 24(23): p. 10277-88. |
| Sharma N, Kumar V, Everingham S, Mali RS, Kapur R, Zeng LF, Zhang ZY, Feng GS, Hartmann K, Roers A, Craig AW. SH2 domain-containing phosphatase 2 is a critical regulator of connective tissue mast cell survival and homeostasis in mice. Mol Cell Biol. 2012 Jul;32(14):2653-63. Epub 2012 May 7. |
| Sperr, W.R. and P. Valent, Diagnosis, progression patterns and prognostication in mastocytosis. Expert Rev Hematol, 2012. 5(3): p. 261-74. |
| Spritz, R.A., et al., Mutations of the KIT (mast/stem cell growth factor receptor) proto-oncogene account for a continuous range of phenotypes in human piebaldism. Am J Hum Genet, 1992. 51(5): p. 1058-65. |
| Takaki, S., et al., Enhanced hematopoiesis by hematopoietic progenitor cells lacking intracellular adaptor protein, Lnk. J Exp Med, 2002. 195(2): p. 151-60. |
| Yang, Y., et al., Pediatric mastocytosis-associated KIT extracellular domain mutations exhibit different functional and signaling properties compared with KIT-phosphotransferase domain mutations. Blood, 2010. 116(7): p. 1114-23. |
| Zeng, S., et al., Regulation of stem cell factor receptor signaling by Cbl family proteins (Cbl-b/c-Cbl). Blood, 2005. 105(1): p. 226-32. |
| Zermati, Y., et al., Effect of tyrosine kinase inhibitor STI571 on the kinase activity of wild-type and various mutated c-kit receptors found in mast cell neoplasms. Oncogene, 2003. 22(5): p. 660-4. |