Introduction
The most striking morphological feature of mast cells is their numerous cytoplasmic secretory granules. These are filled with preformed compounds (“mediators”), which are rapidly released when mast cells are activated in a way that causes their degranulation. The preformed mediators encompass numerous different substances, including biogenic amines (histamine, serotonin, dopamine), preformed cytokines (e.g. TNF), proteoglycans of serglycin type, various lysosomal enzymes (e.g. b-hexosaminidase) as well as a number of mast cell-specific proteases. The latter include serine proteases of tryptase and chymase type, as well as a Zinc-containing metalloprotease of carboxypeptidase A type (CPA3)(1-3). In addition to the mast cell-specific proteases, mast cell granules have been shown to contain a number of other proteases that are not unique to mast cells, such as various cysteine- and aspartic acid cathepsins, matrix metalloproteases (MMPs), renin and cathepsin G (4). However, in the following, the term “mast cell proteases” refers to the mast cell-specific proteases of chymase, tryptase and CPA3 type.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Structure/activity
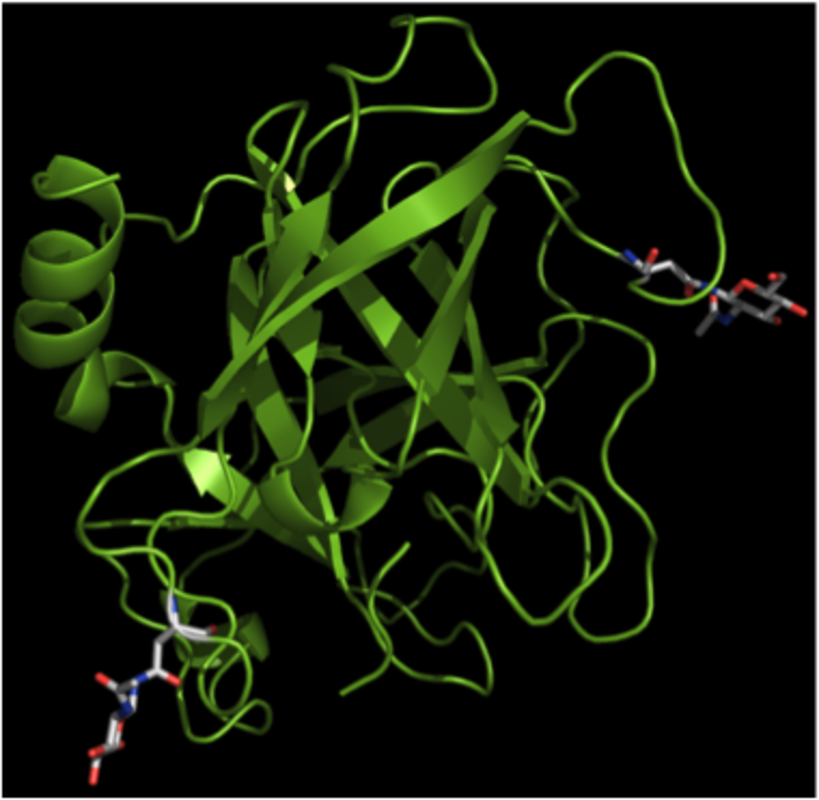
Tryptases have substrate specificities that are reminiscent of that of pancreatic trypsin, i.e., they show a strong preference for cleaving peptide bonds on the C-terminal side of arginine or lysine residues (arginine/lysine in the “P1” position). In analogy, chymases are known to have substrate specificities similar to that of pancreatic chymotrypsin, cleaving peptide bonds preferentially at the C-terminal side of aromatic amino acid residues. Both tryptase and chymases are endopeptidases, i.e. they can cleave internal sites of proteins/peptides. In contrast, CPA3 is an exopeptidase that can only cleave off amino acid residues at the C-terminal end of proteins/peptides, with a preference for aromatic amino acid residues.
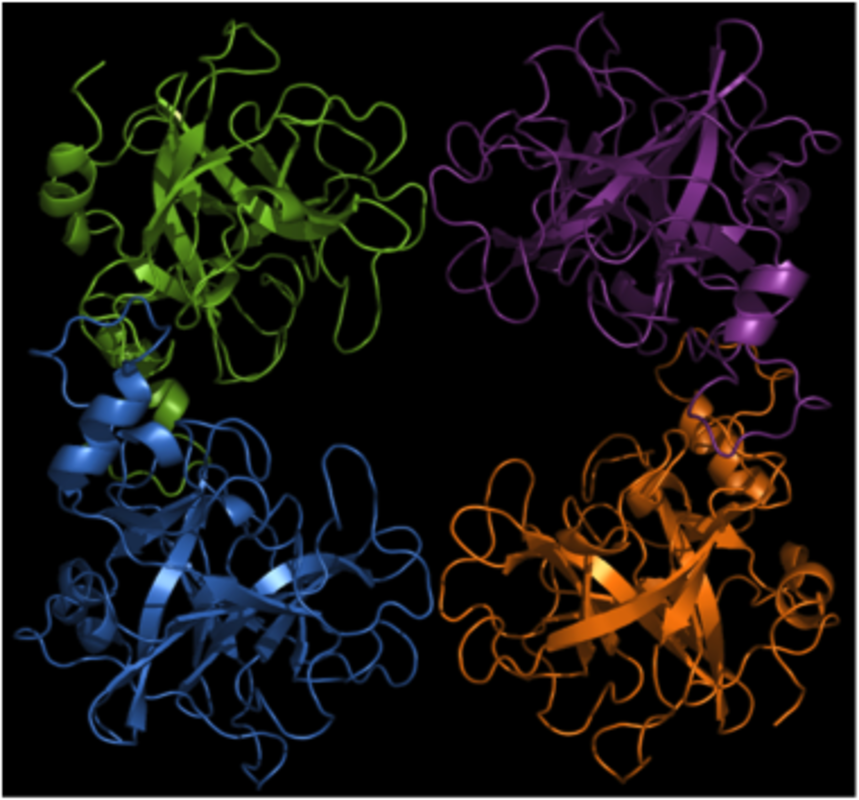
Active tryptase is organized as a tetramer, with all of the active sites facing a narrow central pore (5). This organization restricts the access of macromolecular protease inhibitors as well as of potential substrates. As a result, tryptase is resistant to inhibition by naturally occurring protease inhibitors and has a rather narrow substrate cleavage profile. The three-dimensional structure of chymase has been resolved (6) whereas the structure of CPA3 has not yet been elucidated.
Expression in mast cell subclasses
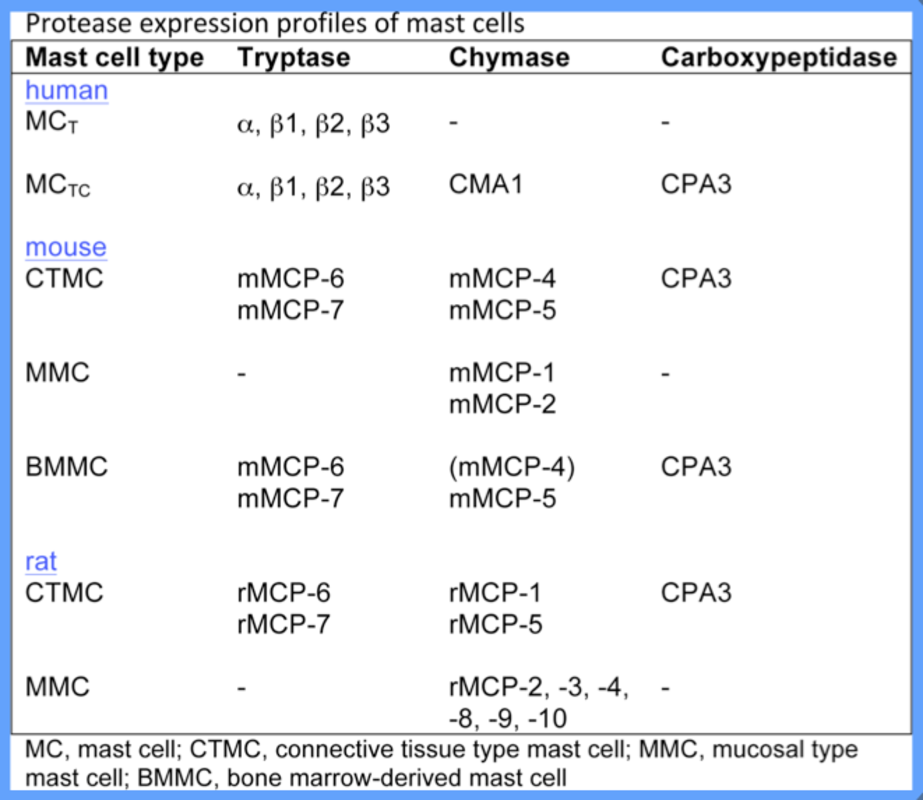
The expression profile of mast cell proteases varies extensively among mast cell subsets. In rodents, mast cells are subdivided into connective tissue type mast cells (CTMCs) and mucosal type mast cells (MMCs), and these are distinguished by their respective protease expression profiles: murine CTMCs express the chymases mouse mast cell protease (mMCP)-4, mMCP-5, the tryptases mMCP-6 and mMCP-7 and CPA3; In contrast, MMCs express the chymases mMCP-1 and mMCP-2 but do not express tryptases or CPA3. In humans, mast cells are classified as either “MCT”, which express tryptase only, or “MCTC”, which express tryptase, chymase and CPA3.
Function
The mast cell proteases are major components of mast cell granules, constituting up to 50% of the total protein content of a mast cell. Thus, when mast cells degranulate, very large amounts of proteases are released into the tissue and these proteases will most likely have a major impact on any condition in which mast cell degranulation occurs. Notably, differently to most other proteases (e.g. pancreatic digestive proteases and MMPs), the mast cell proteases are all stored in fully active form, i.e. with their activation peptides cleaved off. Consequently, the proteases released after mast cell degranulation will have the capacity to immediately carry out proteolytic events in the tissue.
In order to elucidate the function of the mast cell proteases, murine knockout animals have been generated lacking expression of mMCP-1 (7), mMCP-4 (8), mMCP-5 (9), mMCP-6 (10, 11) and CPA3 (12). In addition, a mouse with a mutated active site of CPA3 has been generated (13). These strains have been evaluated in a number of murine disease models, in particular in settings where a global deficiency of mast cells has been shown to have an impact. These investigations have shown that chymase mMCP-4 has harmful roles in a model for abdominal aortic aneurysm formation (14), glomerulonephritis (15), skin blistering (16) and in the inflammatory response following an epidermal burn (9), but also that mMCP-4 has a protective role in allergic airway inflammation (17), towards certain toxins (18) and in sepsis (19). The tryptase mMCP-6 has been shown to have harmful effects in the context of arthritis (20, 21) and experimental colitis (13), while being protective in the context of Trichinella spiralis (10) and Klebisella pneumoniae infection (11). CPA3 has been shown to be essential for protection towards certain snake venom toxins and towards the endogenous toxin endothelin-1 (13, 22).
Altogether, recent studies have shown that mast cell proteases can have a major detrimental impact on a number of settings in which mast cells previously have been implicated. Mast cell proteases are thereby emerging as major effector molecules mediating detrimental effects ascribed to mast cells. Based on these findings, mast cell proteases may accordingly be evaluated as potential drug target candidates in various diseases (2). However, mast cell protease inhibition should be carried out with great caution, as mast cell proteases in several settings have been shown to have protective functions.
References:
| 1. | Pejler, G., Rönnberg, E., Waern, I., and Wernersson, S. (2010) Mast cell proteases: multifaceted regulators of inflammatory disease. Blood 115, 4981-4990 |
| 2. | Pejler, G., Åbrink, M., Ringvall, M., and Wernersson, S. (2007) Mast cell proteases. Adv Immunol 95, 167-255 |
| 3. | Pejler, G., Knight, S. D., Henningsson, F., and Wernersson, S. (2009) Novel insights into the biological function of mast cell carboxypeptidase A. Trends Immunol 30, 401-408 |
| 4. | Lundequist, A., and Pejler, G. (2011) Biological implications of preformed mast cell mediators. Cell Mol Life Sci 68, 965-975 |
| 5. | Pereira, P. J., Bergner, A., Macedo-Ribeiro, S., Huber, R., Matschiner, G., Fritz, H., Sommerhoff, C. P., and Bode, W. (1998) Human beta-tryptase is a ring-like tetramer with active sites facing a central pore. Nature 392, 306-311 |
| 6. | Reiling, K. K., Krucinski, J., Miercke, L. J., Raymond, W. W., Caughey, G. H., and Stroud, R. M. (2003) Structure of human pro-chymase: a model for the activating transition of granule-associated proteases. Biochemistry 42, 2616-2624 |
| 7. | Knight, P. A., Wright, S. H., Lawrence, C. E., Paterson, Y. Y., and Miller, H. R. (2000) Delayed expulsion of the nematode Trichinella spiralis in mice lacking the mucosal mast cell-specific granule chymase, mouse mast cell protease-1. J Exp Med 192, 1849-1856. |
| 8. | Tchougounova, E., Pejler, G., and Abrink, M. (2003) The chymase, mouse mast cell protease 4, constitutes the major chymotrypsin-like activity in peritoneum and ear tissue. A role for mouse mast cell protease 4 in thrombin regulation and fibronectin turnover. J Exp Med 198, 423-431 |
| 9. | Younan, G., Suber, F., Xing, W., Shi, T., Kunori, Y., Abrink, M., Pejler, G., Schlenner, S. M., Rodewald, H. R., Moore, F. D., Jr., Stevens, R. L., Adachi, R., Austen, K. F., and Gurish, M. F. (2010) The inflammatory response after an epidermal burn depends on the activities of mouse mast cell proteases 4 and 5. J Immunol 185, 7681-7690 |
| 10. | Shin, K., Watts, G. F., Oettgen, H. C., Friend, D. S., Pemberton, A. D., Gurish, M. F., and Lee, D. M. (2008) Mouse mast cell tryptase mMCP-6 is a critical link between adaptive and innate immunity in the chronic phase of Trichinella spiralis infection. J Immunol 180, 4885-4891 |
| 11. | Thakurdas, S. M., Melicoff, E., Sansores-Garcia, L., Moreira, D. C., Petrova, Y., Stevens, R. L., and Adachi, R. (2007) The mast cell-restricted tryptase mMCP-6 has a critical immunoprotective role in bacterial infections. J Biol Chem 282, 20809-20815 |
| 12. | Feyerabend, T. B., Hausser, H., Tietz, A., Blum, C., Hellman, L., Straus, A. H., Takahashi, H. K., Morgan, E. S., Dvorak, A. M., Fehling, H. J., and Rodewald, H. R. (2005) Loss of histochemical identity in mast cells lacking carboxypeptidase A. Mol Cell Biol 25, 6199-6210 |
| 13. | Schneider, L. A., Schlenner, S. M., Feyerabend, T. B., Wunderlin, M., and Rodewald, H. R. (2007) Molecular mechanism of mast cell mediated innate defense against endothelin and snake venom sarafotoxin. J Exp Med 204, 2629-2639 |
| 14. | Sun, J., Zhang, J., Lindholt, J. S., Sukhova, G. K., Liu, J., He, A., Abrink, M., Pejler, G., Stevens, R. L., Thompson, R. W., Ennis, T. L., Gurish, M. F., Libby, P., and Shi, G. P. (2009) Critical role of mast cell chymase in mouse abdominal aortic aneurysm formation. Circulation 120, 973-982 |
| 15. | Scandiuzzi, L., Beghdadi, W., Daugas, E., Abrink, M., Tiwari, N., Brochetta, C., Claver, J., Arouche, N., Zang, X., Pretolani, M., Monteiro, R. C., Pejler, G., and Blank, U. (2010) Mouse mast cell protease-4 deteriorates renal function by contributing to inflammation and fibrosis in immune complex-mediated glomerulonephritis. J Immunol 185, 624-633 |
| 16. | Lin, L., Bankaitis, E., Heimbach, L., Li, N., Abrink, M., Pejler, G., An, L., Diaz, L. A., Werb, Z., and Liu, Z. (2011) Dual targets for mouse mast cell protease-4 in mediating tissue damage in experimental bullous pemphigoid. J Biol Chem 286, 37358-37367 |
| 17. | Waern, I., Jonasson, S., Hjoberg, J., Bucht, A., Abrink, M., Pejler, G., and Wernersson, S. (2009) Mouse mast cell protease 4 is the major chymase in murine airways and has a protective role in allergic airway inflammation. J Immunol 183, 6369-6376 |
| 18. | Akahoshi, M., Song, C. H., Piliponsky, A. M., Metz, M., Guzzetta, A., Abrink, M., Schlenner, S. M., Feyerabend, T. B., Rodewald, H. R., Pejler, G., Tsai, M., and Galli, S. J. (2011) Mast cell chymase reduces the toxicity of Gila monster venom, scorpion venom, and vasoactive intestinal polypeptide in mice. J Clin Invest 121, 4180-4191 |
| 19. | Piliponsky, A. M., Chen, C. C., Rios, E. J., Treuting, P. M., Lahiri, A., Abrink, M., Pejler, G., Tsai, M., and Galli, S. J. (2012) The Chymase Mouse Mast Cell Protease 4 Degrades TNF, Limits Inflammation, and Promotes Survival in a Model of Sepsis. Am J Pathol 181, 875-886 |
| 20. | McNeil, H. P., Shin, K., Campbell, I. K., Wicks, I. P., Adachi, R., Lee, D. M., and Stevens, R. L. (2008) The mouse mast cell-restricted tetramer-forming tryptases mouse mast cell protease 6 and mouse mast cell protease 7 are critical mediators in inflammatory arthritis. Arthritis and rheumatism 58, 2338-2346 |
| 21. | Shin, K., Nigrovic, P. A., Crish, J., Boilard, E., McNeil, H. P., Larabee, K. S., Adachi, R., Gurish, M. F., Gobezie, R., Stevens, R. L., and Lee, D. M. (2009) Mast cells contribute to autoimmune inflammatory arthritis via their tryptase/heparin complexes. J Immunol 182, 647-656 |
| 22. | Metz, M., Piliponsky, A. M., Chen, C. C., Lammel, V., Abrink, M., Pejler, G., Tsai, M., and Galli, S. J. (2006) Mast cells can enhance resistance to snake and honeybee venoms. Science 313, 526-530 |