Introduction
A hallmark feature of mast cells from all species is their strong metachromatic staining with various cationic dues, such as Toluidine blue. These properties have been used since the late 19th century to identify mast cells as such (1). The biochemical background for the characteristic staining properties of mast cells is that their secretory granules contain large amounts of proteoglycans.
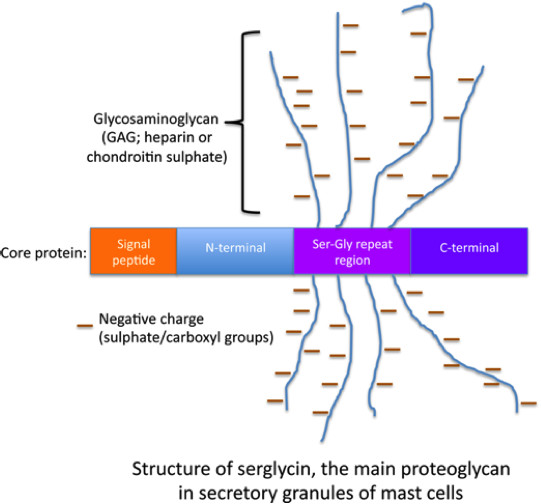{kind=link}
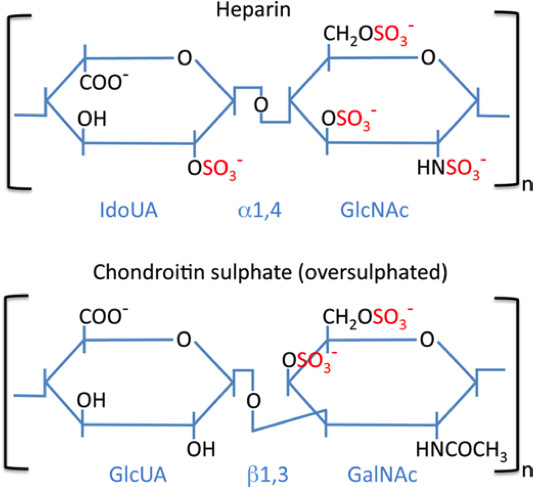{kind=link}
{kind=link}
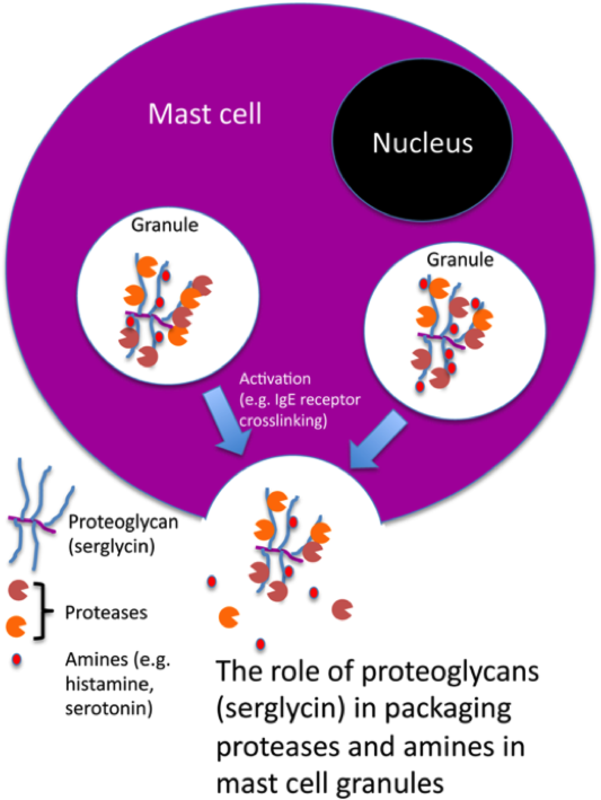{kind=link}
Structure
All proteoglycans are built up by a “core” protein to which unbranched polysaccharide chains of glycosaminoglycan (GAG) type are attached. A large number of different types of proteoglycans are expressed by mammals, all being distinguished by their specific type of core protein but also by the nature of the GAG chains attached (2). In mast cell granules, the dominating (if not only) type of proteoglycan is a species denoted “serglycin” [gene name: SRGN (human), Srgn (mouse)] (3, 4). The serglycin core protein is relatively small (approx. 17 kDa) and its name was coined based on the presence of an extended serine-glycine repeat sequence, in which the serine residues constitute attachment sites for GAGs. The GAGs attached may be of either chondroitin sulphate or heparin type. Chondroitin sulphate is composed of repeating glucuronic acid (GlcUA)-N-acetylgalactosamine (GalNAc) disaccharide units whereas heparin is composed of repeating units of iduronic acid (IdoUA)-N-acetylglucosamine (GlcNAc). Both chondroitin sulphate and heparin contain O-sulphate groups at various positions: chondroitin sulphate at carbon 4 and/or carbon no. 6 of the GalNAc residues; heparin at carbon no. 2 of the IdoUA residues, at carbon 3 (rare substitution) and carbon no. 6 of the GlcNAc units, and most GlcNAc units are also N-sulphated. Importantly, the exact sulphation patterns and chain lengths of GAGs (in all types of proteoglycans) vary to a large extent, hence all GAGs are heterogeneous in structure. A distinguishing feature of serglycin expressed by mast cells (serglycin is also expressed by other cell types), as opposed to all other types of proteoglycans, is its remarkably high extent of overall sulphation, resulting in a very high total anionic charge density. This forms the basis for the strong staining with cationic dyes. Another feature that distinguishes mast cell serglycin from all other proteoglycan species is its strong resistance to degradation by proteases. Most likely, this property arises from the dense substitution of the serglycin core protein with GAGs, with the GAGs sterically blocking the access of proteolytic enzymes.
Proteoglycan expression by mast cell subclasses
Although serglycin is the dominating proteoglycan species in all types of mast cells, the nature of the attached GAG chains varies to a large extent between mast cells of different subclasses. The most notable example of this is the sub-classification of connective tissue type mast cells (CTMCs) and mucosal-type mast cells (MMCs) of rodents, where the proteoglycans in CTMCs (e.g. in skin, peritoneum) contain mostly heparin (attached to the serglycin core protein) whereas chondroitin sulphate is the dominating GAG type attached to the serglycin core protein of MMCs (e.g. in intestinal mucosa) (5).
Function
A breakthrough in the elucidation of the function of mast cell proteoglycans came when the gene coding for N-deacetylase/N-sulphotransferase 2 (NDST-2) was knocked out simultaneously by two groups (6, 7). NDST-2 was shown to be critical for sulphation of the heparin present in CTMCs, whereas GAGs of chondroitin sulphate type are not affected, and it was shown that the absence of functional NDST-2 resulted in an almost complete inability of CTMCs to store a number of mast cell-specific proteases of chymase (mMCP-4, mMCP-5), tryptase (mMCP-6) and carboxypeptidase A (CPA3) type (6, 7). In addition, the storage of histamine was defective. The conclusion from these findings was that the mast cell proteoglycans are essential for promoting the storage of proteases and histamine in granules. Further proof for this notion came when the gene coding for the serglycin core protein was ablated in mice (8, 9). It was shown that the absence of serglycin resulted in a similar phenotype in CTMCs as seen in the absence of NDST-2. Serglycin was also shown to be essential for promoting the storage of granule compounds in mast cell types expressing chondroitin sulphate rather than heparin (e.g. bone marrow derived mast cells, MMCs). Another striking finding was that the absence of serglycin (in all mast cell types) resulted in a completely abrogated metachromatic staining. Hence, the morphological criteria that are routinely used for identification of mast cells are totally dependent on their content of proteoglycans of the serglycin species.
In addition to promoting the actual storage of mast cell proteases, it has been shown that mast cell proteoglycans may influence the activities of these enzymes. A notable example is tryptase, an enzyme that is active as a tetramer. It has been shown that the stability of the tryptase tetramer is dependent on its interaction with heparin, i.e. that tryptase will rapidly dissociate into monomers and thereby loose activity in the absence of heparin. Vice versa, it has been shown that the tetramerization of tryptase, with accompanying gaining of enzymatic activity, requires the presence of heparin that acts by crosslinking the individual tryptase monomers (10). As for chymase, heparin has been shown protect the enzyme from neutralization by different endogenous protease inhibitors, thereby increasing its lifespan following exocytosis from granules. Moreover, the association of chymase with heparin (serglycin) has been shown to promote the degradation of chymase substrates (11).
The dominating fate of the mast cell proteoglycans (serglycin) and its associated compounds is secretion following activation of mast cells in various ways. However, recent findings have shown that mast cell proteoglycans may also leak into the cytosol as a process accompanying cell death. Intriguingly, it has been shown that serglycin (and its associated proteases) thereby promotes apoptotic over necrotic cell death (12).
References:
| 1. | Ehrlich, P. (1878) Beitrage zur Theorie und Praxis der Histologishen Farbung. Thesis, Lepzig |
| 2. | Bishop, J. R., Schuksz, M., and Esko, J. D. (2007) Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 446, 1030-1037 |
| 3. | Kolset, S. O., Pejler, G. (2011) Serglycin- A Structural And Functional Chameleon With Wide Impact On Immune Cells. J. Immunol. 187, 4927-4933 |
| 4. | Rönnberg, E., Melo, F., Pejler, G. (2012) Mast Cell Proteoglycans. J Histochem Cytochem in press |
| 5. | Enerbäck, L., Kolset, S. O., Kusche, M., Hjerpe, A., and Lindahl, U. (1985) Glycosaminoglycans in rat mucosal mast cells. Biochem J 227, 661-668 |
| 6. | Forsberg, E., Pejler, G., Ringvall, M., Lunderius, C., Tomasini-Johansson, B., Kusche-Gullberg, M., Eriksson, I., Ledin, J., Hellman, L., and Kjellén, L. (1999) Abnormal mast cells in mice deficient in a heparin-synthesizing enzyme. Nature 400, 773-776 |
| 7. | Humphries, D. E., Wong, G. W., Friend, D. S., Gurish, M. F., Qiu, W. T., Huang, C., Sharpe, A. H., and Stevens, R. L. (1999) Heparin is essential for the storage of specific granule proteases in mast cells. Nature 400, 769-772 |
| 8. | Åbrink, M., Grujic, M., and Pejler, G. (2004) Serglycin is essential for maturation of mast cell secretory granule. J Biol Chem 279, 40897-40905 |
| 9. | Braga, T., Grujic, M., Lukinius, A., Hellman, L., Åbrink, M., and Pejler, G. (2007) Serglycin proteoglycan is required for secretory granule integrity in mucosal mast cells. Biochem J 403, 49-57 |
| 10. | Hallgren, J., and Pejler, G. (2006) Biology of mast cell tryptase. An inflammatory mediator. Febs J 273, 1871-1895 |
| 11. | Pejler, G., Åbrink, M., Ringvall, M., and Wernersson, S. (2007) Mast cell proteases. Adv Immunol 95, 167-255 |
| 12. | Melo, F. R., Grujic, M., Spirkoski, J., Calounova, G., and Pejler, G. (2012) Serglycin proteoglycan promotes apoptotic versus necrotic cell death in mast cells. J Biol Chem 287, 18142-18152 |