Ca2+ entry pathways for mast cell activation
In mast cells, the increase of free cytosolic Ca2+ regulates a variety of cellular processes including degranulation with release of preformed inflammatory mediators, production of eicosanoids, activation of transcription factors including the nuclear factor of activated T cells (NFAT) and synthesis of cytokines as well as migration and chemotaxis (1). The importance of extracellular Ca2+ as a requirement for anaphylactic release of histamine from rat peritoneal mast cells was already shown in 1972 (1) leading to the concept that antigen-mediated mast cells degranulation is dependent on Ca2+ influx through Ca2+ channels (1). Antigen-induced aggregation of high-affinity receptors for IgE (FcεRI) activates a large number of signalling molecules (2). A major downstream target is phospholipase Cγ1 (PLCγ1), which catalyses formation of IP3 and release of Ca2+ from intracellular stores, followed by an influx of Ca2+ from the extracellular space (capacitative or store-operated Ca2+ entry (CCE, SOCE)) (3, 4). Ionic currents mediating this Ca2+ influx in mast cells were first characterised in RBL cells and rat peritoneal mast cells (5). Recently, Orai1 was identified as the pore forming CRAC channel subunit. SOCE is substantially decreased in Orai1-deficient BMMCs (6), and the FcεRI-mediated Ca2+ entry is reduced by 70%, while the cells completely lack detectable Ca2+ (CRAC) currents (7). This suggests that other channel proteins including Orai2, Orai3 and/or members of the TRP family such as TRPC1 (8) and TRPC5 (9) might also contribute to Ca2+ entry (10).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cation channels formed by TRP proteins
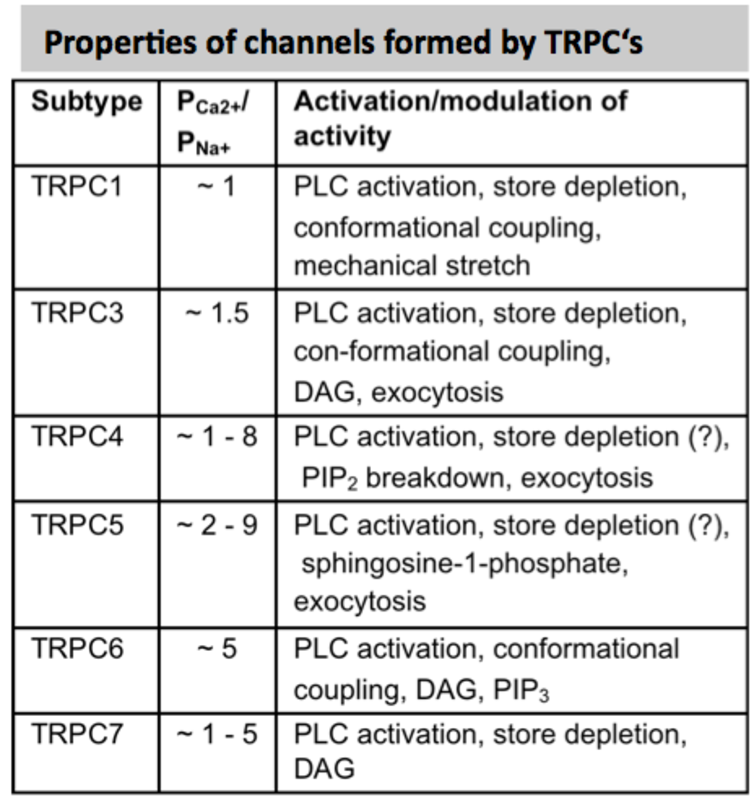
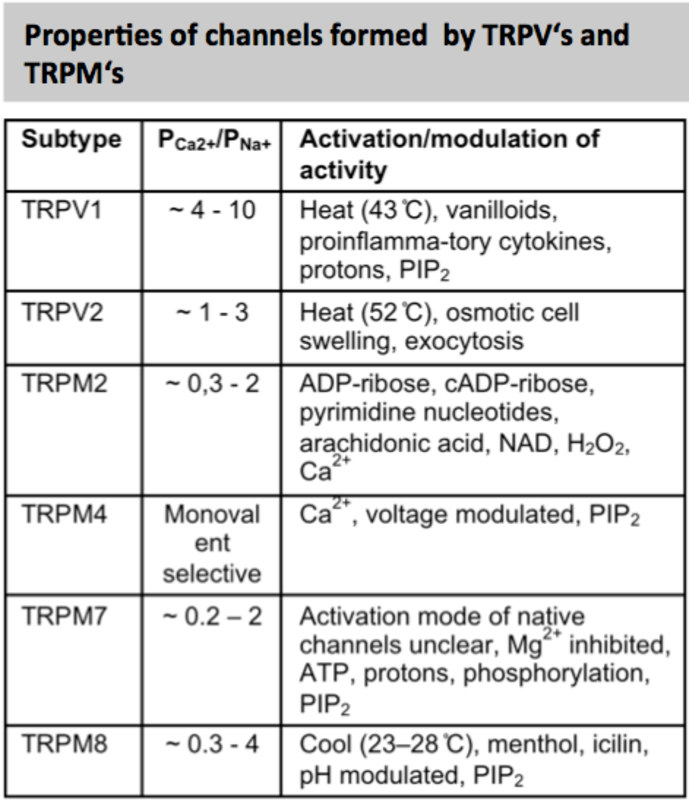
Transient receptor potential (TRP) channels are a large and functionally heterogeneous family of cation-conducting channel proteins, which are expressed in many (if not in all) excitable and non-excitable cells and are involved in sensing of a variety of environmental stimuli such as temperature, pH, osmolarity, pheromones, taste and plant compounds, participate in numerous cellular processes and several mutations in Trp genes appear to be causative factors in rare heritable channelopathies (11). The mammalian 28 TRP proteins are classified according to structural homology into 6 subfamilies: TRPC (canonical), TRPV (vanilloid), TRPM (melastatin), TRPA (ankyrin), TRPML (mucolipin), and TRPP (polycystin) (12, 13). Most TRP channels show little voltage dependence and are nonselective with a permeability for Ca2+ over Na+ (ratio PCa/PNa) below 10 (see panels).
How TRP channels regulate [Ca2+]i and mast cell activation
TRP channels were found to influence cellular Ca2+ signaling by several mechanisms. First, by conducting Ca2+ ions to various degrees TRP channels directly contribute to Ca2+ influx via the plasma membrane (e.g. TRPC1 (8) or TRPC5 (9)). Second, by conducting Na+ TRP channels mediate electrogenic effects through plasma membrane depolarization which may have opposite consequence in different cell types: in contrast to excitable cells, where TRP-mediated Na+ entry and depolarization can enhance Ca2+ entry by gating of voltage-operated Ca2+ channels, they can do the opposite in nonexcitable cells such as mast cells where depolarization reduces the driving force for Ca2+ entry through CRAC channels (e.g. TRPM4, see ref. 14). In addition, there is emerging evidence that several TRP channels are also located in the membrane of the sarco/endoplasmic reticulum (SR/ER), endosomes, lysosomes or other intracellular vesicles where they can serve as Ca2+ release channels and conduct Ca2+ from the luminal stores into the cytoplasm (for more details see (15)). The relevance of such channels for specific mast cell functions is still elusive.
TRPs in mast cells: expression pattern and postulated functions
TRPC channels: The mammalian members of the TRPC family can be divided into 3 subfamilies on the basis of functional similarities and sequence homology: TRPC1/TRPC4/TRPC5, TRPC3/TRPC6/TRPC7 and TRPC2. All TRPC proteins seem to be activated by stimulation of G-protein-coupled receptors or receptor tyrosine kinases. TRPC proteins were reported to form heteromeric channels. TRPC3, TRPC6 and TRPC7 when expressed in heterologous systems are potentiated by direct application of diacylglycerol (DAG) analogs.
TRPC1 has been detected in murine BMMC on mRNA (8) and protein (16) level and using a microarray expression analysis in human skin mast cells (17). Impaired degranulation of BMMCs induced by down-regulation of TRPC1 expression using siRNAs could be rescued by exogenous expression of TRPC1 (8). Expression of TRPC3 was reported in mouse BMMC via Western Blot (16) and RT-PCR. In BMMCs, also transcripts of Trpc2 (9), transcripts of Trpc4 and Trpc5 (8) and TRPC4 proteins (16) were found. Trpc6 and Trpc7 transcripts and proteins were also found in mouse BMMC (16) and based on immunoprecipitation experiments using an antibody that was designed to detect all three members of the TRPC3/TRPC6/TRPC7 subfamily. The generation and use of antibodies that specifically detect individual TRP proteins is an critical issue in TRP channel research similarly like for studies on other membrane proteins (see ref. 18)
TRPV channels: Similar to the TRPC family, the TRPV (vanilloid) family can be divided into two subfamilies on the basis of structure, function and Ca2+ selectivity: TRPV1-4, and TRPV5/6. In the TRPV subfamily, TRPV1, TRPV2, TRPV3 and TRPV4 are nonselective cation channels that are activated by a different range in temperatures and by numerous chemical stimuli.
TRPV1 proteins are expressed in mast cells of the human bladder (19) and skin (20). The TRPV1 specific agonists capsaicin or resiniferatoxin induced calcium uptake in several mouse mast cell lines, in BMMC, but not in PMC (21).
TRPM channels: The members of the TRPM (melastatin) family are divided into 4 groups: TRPM1/3, TRPM2/8, TRPM4/5, and TRPM 6/7. TRPM2 is activated by ADP ribose (EC50, 100 µM) and activated by H2O2 and under conditions of ROS production, TRPM4 and TRPM5 are the only monovalent-selective ion channels of the TRP family and are activated by an increase in Ca2+ levels in the cytosol.
TRPM6 and TRPM7 proteins form Mg2+ and Ca2+ conducting channels (inhibited by intracellular Mg2) and possess also protein kinase activity. TRPM7 is ubiquitously expressed and deletion of the Trpm7 gene leads to embryonic lethality. TRPM8 can be activated by cold (8–28°C) and enhanced by cooling compounds such as menthol and icilin.
Trpm2 transcripts were identified in human lung mast cells and cord blood derived mast cells in a microarray expression analysis (17). In mouse BMMC, Trpm4 transcripts were detected using Northern blot analysis and the 138 kDa TRPM4 proteins were identified in BMMCs of wild type but not of Trpm4-/- mice (14). It could be shown that TRPM4 channels depolarize the membrane following adenosine- and FcεRI-stimulation and, thereby, critically decrease the Ca2+ influx in BMMC’s via CRAC channels (see model in Fig. 1). Accordingly, activated Trpm4-/- BMMCs release excessive histamine, leukotrienes and tumor necrosis factor (TNF), and Trpm4-/- animals display a more severe acute anaphylactic response in the skin compared to wild-type controls (14). Expression of TRPM7 has been found in human lung mast cells (22) and Mg2+-inhibited currents (MIC), which can be mediated by TRPM7 proteins, were identified in human lung mast cells. Downregulation of TRPM7 expression in human mast lung cells resulted in significant reduction of MIC currents and mast cell survival which could not be rescued by an increase in the extracellular Mg2+ concentration (22). Trpm7-deficient mice as well as mice homozygous for an allele lacking the kinase domaine of TRPM7 (Trpm7Δkinase/Δkinase) die early during development (23), but Trpm7+/Δkinase are viable and MIC currents are significantly reduced in PMCs of these mice (23). TRPM8 proteins have also been proposed as mediators of mast cell activation e.g. in the development of cold urticaria. However, Medic et al. found no deficits in mast cell activation using Ca2+ imaging, mast cell degranulation and development of passive anaphylaxis in Trpm8-/- mice, which makes a role of TRPM8 for mast cell activation in cold urticaria unlikely (24).
References:
| 1. | Ma HT & Beaven MA (2009) Crit Rev Immunol 29, 155-186. |
| 2. | Blank U & Rivera J (2004) Trends Immunol 25, 266-273. |
| 3. | Parekh AB (2010) Nat Rev Drug Discov 9, 399-410. |
| 4. | Putney JW, Jr. (1986) Cell Calcium 7, 1-12. |
| 5. | Hoth M & Penner R (1992) Nature 355, 353-356. |
| 6. | Gwack Y, Srikanth S, Oh-Hora M, Hogan PG, Lamperti ED, Yamashita M, Gelinas C, Neems DS, Sasaki Y, Feske S, et al. (2008) Mol Cell Biol 28, 5209-5222. |
| 7. | Vig M, DeHaven WI, Bird GS, Billingsley JM, Wang H, Rao PE, Hutchings AB, Jouvin MH, Putney JW, & Kinet JP (2008) Nat Immunol 9, 89-96. |
| 8. | Suzuki R, Liu X, Olivera A, Aguiniga L, Yamashita Y, Blank U, Ambudkar I, & Rivera J (2010) J Leukoc Biol 88, 863-875. |
| 9. | Ma HT, Peng Z, Hiragun T, Iwaki S, Gilfillan AM, & Beaven MA (2008) J Immunol 180, 2233-2239. |
| 10. | Freichel M, Almering J, & Tsvilovskyy V Front Immunol 3, 150. |
| 11. | Nilius B & Owsianik G (2010) Pflugers Arch 460, 437-450. |
| 12. | Clapham DE, Julius D, Montell C, & Schultz G (2005) Pharmacol Rev 57, 427-450. |
| 13. | Montell C, Birnbaumer L, & Flockerzi V (2002) Cell 108, 595-598. |
| 14. | Vennekens R, Olausson J, Meissner M, Bloch W, Mathar I, Philipp SE, Schmitz F, Weissgerber P, Nilius B, Flockerzi V, et al. (2007) Nat Immunol 8, 312-320. |
| 15. | Dong XP, Wang X, & Xu H (2010) J Neurochem 113, 313-328. |
| 16. | Hernandez-Hansen V, Smith AJ, Surviladze Z, Chigaev A, Mazel T, Kalesnikoff J, Lowell CA, Krystal G, Sklar LA, Wilson BS, et al. (2004) J Immunol 173, 100-112. |
| 17. | Bradding P, Okayama Y, Kambe N, & Saito H (2003) J Leukoc Biol 73, 614-620. |
| 18. | Meissner M, Obmann V, Hoschke M, Link S, Jung M, Held G, Philipp S, Zimmermann R, & Flockerzi V (2011) in TRP Channels, ed. Zhu M (CRC Press ). |
| 19. | Lazzeri M, Vannucchi MG, Zardo C, Spinelli M, Beneforti P, Turini D, & Faussone-Pellegrini MS (2004) Eur Urol 46, 792-798. |
| 20. | Stander S, Moormann C, Schumacher M, Buddenkotte J, Artuc M, Shpacovitch V, Brzoska T, Lippert U, Henz BM, Luger TA, et al. (2004) Exp Dermatol 13, 129-139. |
| 21. | Biro T, Maurer M, Modarres S, Lewin NE, Brodie C, Acs G, Acs P, Paus R, & Blumberg PM (1998) Blood 91, 1332-1340. |
| 22. | Wykes RC, Lee M, Duffy SM, Yang W, Seward EP, & Bradding P (2007) J Immunol 179, 4045-4052. |
| 23. | Ryazanova LV, Rondon LJ, Zierler S, Hu Z, Galli J, Yamaguchi TP, Mazur A, Fleig A, & Ryazanov AG (2010) Nat Commun 1, 109. |
| 24. | Medic N, Desai A, Komarow H, Burch LH, Bandara G, Beaven MA, Metcalfe DD, & Gilfillan AM (2011) Cell Calcium 50, 473-480. |
| 25. | Shumilina E, Lam RS, Wolbing F, Matzner N, Zemtsova IM, Sobiesiak M, Mahmud H, Sausbier U, Biedermann T, Ruth P, et al. (2008) J Immunol 180, 8040-8047. |