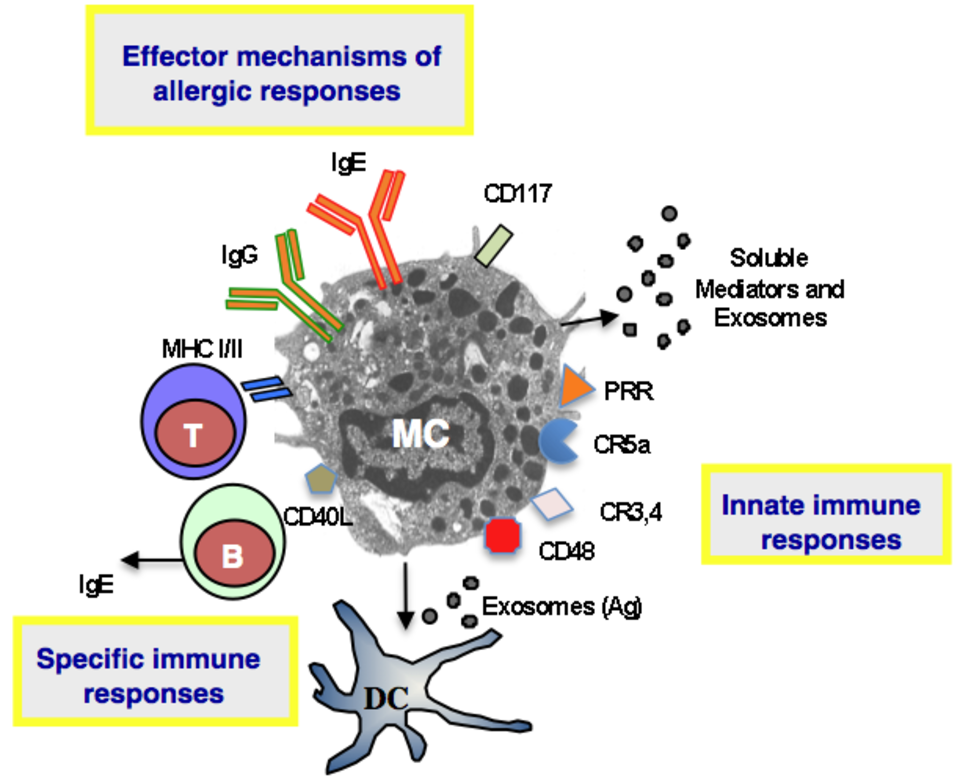
Mast cells (MCs) are now regarded not only as effector cells playing a detrimental role in IgE-mediated allergic responses (Fig. 1) but also and more importantly as key players in host defences including via allergy-mediated processes. In this respect, mast cell responses during hypersensitivity reactions appear to be primarily devoted to anticipate aggressive and noxious environmental stimuli against the host.
Role of MC in innate immunity
The preferential localisation of MCs in close proximity to epithelial layers such as in
the skin, in the respiratory and in the gastrointestinal tracts, predispose these cells to enter into contact with noxious or infectious agents. In this regard, MCs are equipped with a wide range of receptors and pattern recognition molecules to sense their environment. We will present two examples, anti-parasite and anti-bacterial immunity.
Immunity against parasites: although MCs have been globally associated with immunity against parasitic infections, there are however substantial differences considering immunity against various gastrointestinal helminth species. Typically, genetically mast cell-deficient (W/Wv, Sl/Sld) mice or (Ws/Ws) rats exhibit little or no impairment in their ability to expel a primary infection with Nippostrongylus brasiliensis (1, 2), suggesting that MCs are not essential for the host immune response to this nematode. In contrast, intestinal mastocytosis normally observed following Trichinella spiralis infection, which is dependent on SCF, was markedly reduced following in vivo administration of an anti-(c-kit) receptor antibody, and treated mice did not show any evidence of worm expulsion from the gut, suggesting a protective effect of MC in anti T. spiralis immunity (3). More recently MCs have been assigned a critical role in orchestrating type 2 immunity to helminths (4). Indeed, MC-deficient mice and mice treated with the MC stabilizing agent sodium cromoglycate had dramatically reduced type 2 cytokine production and harbored increased parasite burdens following infection with gastrointestinal helminths (Heligmosomoides polygyrus bakeri and Trichuris muris). These IgE-independent innate defence mechanisms operated by MCs were mediated by IL-25, IL-33, and thymic stromal lymphopoietin (TSLP).
Immunity against bacterial infections: two pioneering reports in 1996 by Echtenacher et al. (5) and by Malaviya et al. (6), using two different mouse model systems, described for the first time a major role of mast cells in host defence against bacterial infections. Many other studies have followed which aim was to identify the mechanisms by which MC exert their effector function against pathogens.
TNF-α produced by MC was first identified as the major cytokine that mediate bacterial clearance, but this finding was challenged afterwards. Indeed, while MC failed to protect mice undergoing severe coecal ligation and puncture (CLP), in moderate sepsis induced by CLP, MC clearly mediated protection in reconstituted MC-deficient W/Wv and Wsh/Wsh mice. In this setting, protection was TNFα-independent, as reconstitution of Wsh/Wsh mice with either WT or TNF-deficient MC was equally effective in restoring protection (7). To sense the presence of bacteria in their environment, MC are equipped with a variety of innate immune receptors which interact with bacteria in both direct (opsonin-independent) (Toll-like receptors TLR 2, TLR4, and TLR9, CD48, and nucleotide-binding oligomerization (NOD) proteins) and indirect (opsonin-dependent) (Fcg receptors and complement receptors) mechanisms (8-10) (Fig. 1). CD48 for example, can detect the presence of fimbriated Escherichia coli, Mycobacterium tuberculosis and Staphylococcus aureus (9). As an alternative mean by which MC can also kill bacteria, is the production of antimicrobial peptides. Human as well as mouse MC have been shown to express and produce cathelicidins and a beta-defensin, and MCs derived from cathelin-related AMP gene (Cnlp-/-) animals had a 50% reduction in their ability to kill group A Streptococcus (11).
Role of MC in adaptive immunity
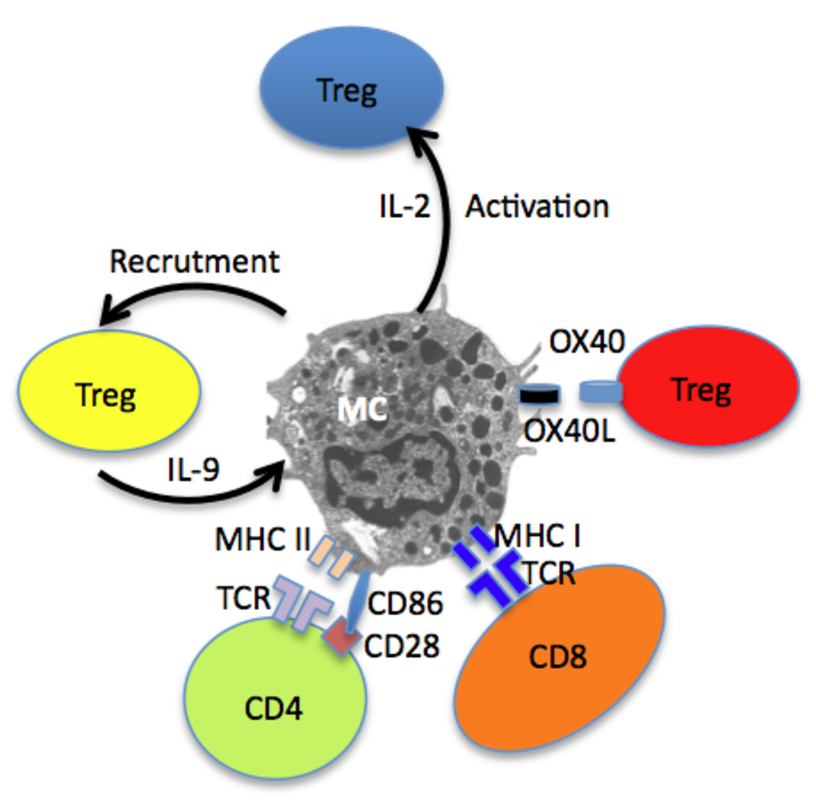
Besides their contribution to host defence via innate mechanisms, MC also promote adaptive immune responses through physical interactions with CD4 and CD8 T cells (Fig. 2). It has been clearly shown that MCs can be primed to express functional MHC class II and costimulatory molecules and can serve as antigen presenting cells for CD4+ T lymphocytes. Under certain conditions, notably exposure to IFN-γ, bone marrow derived MC (BMMC) can be induced to express MHC II molecules (12). This expression occurs not only in mice but also in humans as a small fraction of MCs expresses MHC class II molecules in tissues, namely 10–20% in normal human lung, ~7% in normal human skin and ~10% in pleural cavity, and this fraction increases in tuberculin reactive skin and in autoimmune thyroiditis (13). The MHC class II biosynthetic pathway is peculiar in MC as the bulk of MHC II molecules is found intracellularly and only a minor fraction was expressed at the cell surface of BMMC (14). This is probably the reason why the detection of MHC II molecules on the surface of MC remained elusive for many decades. It was speculated that in immature MCs cathepsin S is expressed at low levels and that appropriate stimuli (such as IFN-γ) enhance cathepsin S expression in MCs allowing maturation of MHC-class II molecules. In BMMC, the bulk of MHC class II molecules is contained in two distinct compartments, with features of both lysosomal compartments and secretory granules defined by their protein content and their accessibility to endocytic tracers. In type I and type II granules, MHC class II molecules, mannose-6-phosphate receptors and lysosomal membrane proteins (lamp1 and lamp2) localize to small intralumenal vesicles. These 60–80-nm vesicles termed exosomes are released along with inflammatory mediators during MC degranulation triggered by IgE-antigen complexes (14) (Fig. 1). Interestingly, when fed with exogenous antigens, MCs can release these exosomes that harbour antigen-derived peptides. These antigen-containing exosomes can stimulate the maturation of mouse dendritic cells leading to enhanced antigen presentation to T cells (15) (Fig. 1). They were also found to be responsible for MC-driven B cell proliferation and activation (16). Interestingly, it has been shown that important co-stimulatory molecules, such as MHC-II, CD86, CD40, CD40L, LFA-1, and ICAM-1, were associated with exosomes (17).
In addition to CD4+ T cells, MCs have been indeed shown to interact with and present antigen also to different T cell subsets among which Treg and CD8+ T lymphocytes (Fig. 2). Similarly, MCs are capable of inducing antigen-specific CD8+ T cell responses in vitro and in vivo. Murine BMMCs can, in fact, process Ag from phagocytosed bacteria for presentation via MHC class I molecules to T cells (18). Moreover, MHC class I-mediated cross presentation of BMMCs and peritoneal MCs to CD8+ T cells was shown to increase CD8+ T cell proliferation, and cytotoxic activity. In turn, CD8+ T cells induce MHC class I and 4-1BB expression on BMMCs as well as the secretion of osteopontin (19). An interesting feature of MCs is their ability to coordinate regulatory response against neoplastic cells, such as hepatocarcinoma in mice, as MCs were found to mobilize myeloid-derived suppressor cells and Treg cells in tumor microenvironment via IL-17 pathway (20).
References:
| 1. | Reed N. D: Function and regulation of mast cells in parasite infections, in Galli SJ, Austen KF (eds): Mast Cell and Basophil Differentiation and Function in Health and Disease. New York, NY, Raven, 1989, p 205 |
| 2. | Arizono N, Kasugai T, Yamada M, Okada M, Morimoto M, Tei H, Newlands GF, Miller HR, Kitamura Y. 1993. Infection of Nippostrongylus brasiliensis induces development of mucosal-type but not connective tissue-type mast cells in genetically mast cell-deficient Ws/Ws rats. Blood 81:2S72, 1993. |
| 3. | Grencis R.K, Else K., Huntley J.F., Nishikawa S. I. 1993. The in vivo role of stem cell factor (c-kit ligand) on mastocytosis and host protective immunity to the intestinal nematode Trichinella spiralis in mice. Parasite Immunol. 1:55-59. |
| 4. | Hepworth M. R., Daniliwicz-Luebert E., Rausch S., Metz M., Klotz C., Maurer M., Hartmann S. 2012. Mast cells orchestrate type 2 immunity to helminths through regulation of tissue-derived cytokines. Proc Natl Acad Sci USA 109 (17):6644-9 |
| 5. | Echtenacher, B., Mannel, D. N., and Hultner, L. 1996. Critical protective role of mast cells in a model of acute septic peritonitis. Nature, 381, 75-77 |
| 6. | Malaviya R, Ikeda T, Ross E, Abraham SN. 1996. Mast cell modulation of neutrophil influx and bacterial clearance at sites of infection through TNF-alpha. Nature 381(6577):77-80. |
| 7. | Piliponsky A. M., Chen C. C., Grimbaldeston M. A., Burns-Guydish S. M., Hardy J., Kalesnikoff J., Contag C. H., Tsai M., Galli S. J. 2010. Mast cell-derived TNF can exacerbate mortality during severe bacterial infections in C57BL/6-KitW-sh/W-sh mice. Am J Pathol 176(2):926-38. |
| 8. | Marshall, J. S. 2004. Mast-cell responses to pathogens. Nat. Rev. Immunol. 4, 787–799. |
| 9. | Abraham S.N, and St. John A. L. 2010. Mast cell-orchestrated immunity to pathogens. Nature Reviews Immunology 10, 440-452 |
| 10. | Ali, H. 2010. Regulation of human mast cell and basophil function by anaphylatoxins C3a and C5a. Immunol. Lett. 128, 36–45. |
| 11. | Di Nardo A., Vitiello A., Gallo R. L. 2003. Cutting edge: mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J Immunol 170(5):2274-8 |
| 12. | Frandji P, Oskéritzian C, Cacaraci F, Lapeyre J, Peronet R, David B, Guillet J. G, Mécheri S. 1993. Antigen-dependent stimulation by bone marrow-derived mast cells of MHC class II-restricted T cell hybridoma. J Immunol. 151(11):6318-28 |
| 13. | Banovac K, Ghandur-Mnaymneh L, Leone J, Neylan D, Rabinovitch A. 1989. Intrathyroidal mast cells express major histocompatibility complex class-II antigens. Int Arch Allergy Appl Immunol. 90(1):43-6. |
| 14. | Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, and Desaymard C. 1997. Accumulation of Major Histocompatibility Complex Class II Molecules in Mast Cell Secretory Granules and Their Release upon Degranulation. Molecular Biology of the Cell 8, 2631–2645. |
| 15. | Skokos D, Goubran Botros H, Demeure C, Morin J, Peronet R, Birkenmeier G, et al. 2003. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J Immunol. 170: 3037–3045. |
| 16. | Tkaczyk, C., Frandji, P., Botros, H. G., Poncet, P., Lapeyre, J., Peronet, R., David, B., and Mecheri, S. 1996. Mouse bone marrow-derived mast cells and mast cell lines constitutively produce B cell growth and differentiation activities. J. Immunol. 157, 1720-1728 |
| 17. | Skokos,D.,Goubran-Botros,H., Roa,M.,and Mecheri,S. 2002. Immunoregulatory properties of mast cell-derived exosomes. Mol. Immunol. 38, 1359-1362 |
| 18. | Malaviya, R., Twesten, N. J., Ross, E. A., Abraham, S. N., and Pfeifer, J. D. 1996. Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells. J. Immunol. 156, 1490-1496. |
| 19. | Stelekati E, Bahri R, D’Orlando O, Orinska Z, Mittrücker HW, Langenhaun R, Glatzel M, Bollinger A, Paus R, Bulfone-Paus S. 2009. Mast cell-mediated antigen presentation regulates CD8+ T cell effector functions. Immunity. 16;31(4):665-76. |
| 20. | Yang Z, Zhang B, Li D, Lv M, Huang C, Shen G-X, et al. 2010. Mast cells mobilize myeloid-derived suppressor cells and Treg cells in tumor microenvironment via IL-17 pathway in murine hepatocarcinoma model. PLoS ONE ; 5: e8922. |

{kind=link}
{kind=link}