Basophils from both human and mouse are effector cells of allergic inflammation. In the last few years it was demonstrated that basophils own additional attributes that enable them to play multifaceted roles in immunoregulation. Namely, it has been shown that basophils initiate Th2 differentiation1-5, regulate plasma cell survival6 and memory7, and enhance the development of autoimmunity such as systemic lupus erythematosus (SLE)8. However, most of these additional attributes have only been shown in the mouse but not in the human system.
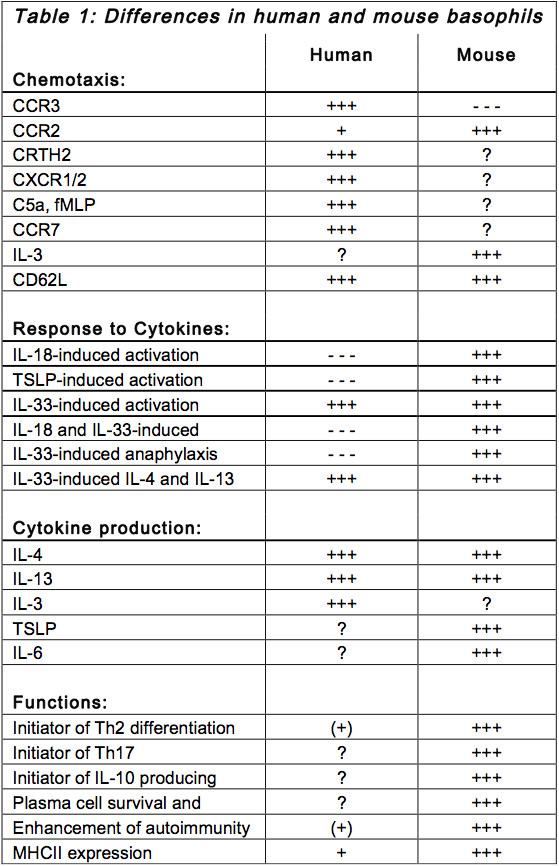
Basophil migration to lymph node and to inflamed tissue and their responses to cytokines are critical in the processes of immunoregulation. Hence this text and table 1 outline some aspects of chemotaxis and cytokine networks of human and mouse basophils. The effects of IL-3, which plays a major role in basophil biology, are summarized in table 2.
{kind=link}
{kind=link}
Chemotaxis
Migration to inflamed tissue
CCR3: Human basophils constitutively express CCR3, which binds besides others eotaxin. Migration of human basophils to inflamed tissue is mainly regulated by eotaxin9. Mouse basophils do not express CCR3 and thus do not respond to eotaxin-induced chemotaxis.
CCR2: On the other hand, CCR2 guides circulation of mouse basophils. But it rather induces mediator release than chemotaxis in human basophils10.
CRTH2: CRTH2, the receptor for prostaglandin D2 (PGD2), has been shown to regulate chemotaxis of both human and mouse basophils11,12. However, while the expression of CRTH2 on human basophils has been clearly shown, its expression on mouse basophils has not been demonstrated so far. Thus it is possible that in humans PGD2 directly attracts CRTH2-positive basophils while in mice other CRTH2-positiv cells (e.g. eosinophils, Th2 cells) may attract basophils via another chemokine such as CCL2.
CXCR1, CXCR2: It has been shown that CXCR1 and CXCR2 regulate human basophil migration via binding of IL-89. Such an effect of CXCR1 and CXCR2 is not known in the mouse system.
Polypetides and lipids: It has been demonstrated that polypeptides and lipids, like C5a and fMLP, serve as chemotactic ligands for human basophils13. Whether C5a and fMLP serve as chemoattractants in mice is not known.
Migration to lymph nodes
CCR7: It has been demonstrated that human basophils express CCR7. Most importantly, adhesion and rolling of human basophils on IL-3-treated human endothelial cells is dependent on CCR7 expression14. Thus the expression of CCR7 potentially regulates migration of human basophils to lymph nodes and may be relevant for the suggested role of basophils in initiatingTh2 differentiation.
IL-3: IL-3 has been shown to be important in regulating basophil migration to draining lymph nodes following helminth infection in mice. However the same study demonstrates that basophils are not necessary for initiating Th2 differentiation15.
CD62L: Since a long time it is clear that expression of CD62L on lymphocytes regulates cell migration to lymph nodes. Thus it is very interesting that activated basophils of Lyn-/- mice and humans suffering from SLE express CD62L8
Cytokine networks
TSLP: Both, human and mouse basophils produce IL-4 and IL-13 upon FceR-crosslinking or IL-3 treatment. Thus it is certain that both, human and mouse basophils support development of Th2 immunity. It has been shown that mouse basophils are capable to initiate Th2 differentiation, too1-5. This capability of basophil-mediated initiation of Th2 differentiation depends however on the type of allergen which was used to induce Th2 immunity. Nevertheless it has been shown that mouse basophils produce thymic stromal lymphopoietin (TSLP) in the lymph nodes and that this TSLP production is crucial for basophil-mediated Th2 differentiation2. But up to now it is not described that human basophils produce TSLP. Furthermore, in mice TSLP promotes differentiation of basophils that show distinct attributes than IL-3-induced basophils16. Although human basophils express the receptor for TSLP effects of TSLP on human basophils are still unknown. Thus it remains uncertain whether and by which mechanism human basophils respond to TSLP and/or initiate Th2 differentiation.
IL-6: Mouse basophils produce IL-6 upon allergen challenge. IL-6 regulates together with IL-4 the survival of plasma cells7 and the induction of IL-10-producing CD8 T cells17. Whether human basophils produce IL-6 is not reported.
IL-33: IL-33 induces IL-4 and IL-13 secretion in both human and mouse basophils. However, IL-33 does not induce histamine release and LTC4 formation by itself in human basophils18. This is in contrast to mouse studies showing that IL-33 induces anaphylaxis independent of antigen challenge but dependent on high IgE levels19.
IL-18: IL-18 activates mouse basophils only. This is peculiar as human basophils constitutively express the IL-18 receptor18. The ligand for the human IL-18 receptor is still not known.
IL-3: IL-3 is able to promote in human basophils the late priming, which occurs as a second phase after prolonged culturing of basophils with IL-3. This IL-3-induced late priming maintains basophils sensitive to C5a triggered LTC4 formation at later time points and transforms them into a state of hyper-responsiveness20. However, it is not clear whether IL-3 promotes a late priming in mice as well. IL-3 is the most potent known cytokine promoting survival of human and mouse basophils in vitro. In humans the IL-3-induced survival is regulated by the expression of Pim-121. The expression of Pim-1 upon IL-3 treatment has not yet been investigated in mouse basophils. Nevertheless it has been shown that IL-3 treatment in vivo results in basophilia in mice22. Furthermore, IL-3 induces in human basophils in addition to Pim-1 de-novo expression of retinaldehyde dehydrogenase-II (RALDH2)23, granzyme B (GzmB)24 and the Aryl-hydrocarbon receptor (AhR). These proteins seem to define the immunoregulatory role of human basophils: Th2 differentiation (RALDH2), cell cytotoxicity (GzmB) and regulation of continuous LTC4 formation and IL-4/IL-13 secretion (AhR together with C5a, own observation). The expression of these proteins has not been investigated in mice so far. Interestingly, human basophils produce IL-3 themselves, indicating that they regulate their function partly in an autocrine fashion25.
References:
| 1. | Oh, K., Shen, T., Le Gros, G. & Min, B. Induction of Th2 type immunity in a mouse system reveals a novel immunoregulatory role of basophils. Blood 109, 2921–2927 (2007). |
| 2. | Sokol, C. L., Barton, G. M., Farr, A. G. & Medzhitov, R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat Immunol 9, 310–318 (2007). |
| 3. | Sokol, C. L. et al. Basophils function as antigen-presenting cells for an allergen-induced T helper type 2 response. Nat Immunol 10, 713–720 (2009). |
| 4. | Perrigoue, J. G. et al. MHC class II-dependent basophil-CD4+ T cell interactions promote T(H)2 cytokine-dependent immunity. Nat Immunol 10, 697–705 (2009). |
| 5. | Yoshimoto, T. et al. Basophils contribute to T(H)2-IgE responses in vivo via IL-4 production and presentation of peptide-MHC class II complexes to CD4+ T cells. Nat Immunol 10, 706–712 (2009). |
| 6. | Rodriguez Gomez, M. et al. Basophils support the survival of plasma cells in mice. The Journal of Immunology 185, 7180–7185 (2010). |
| 7. | Denzel, A. et al. Basophils enhance immunological memory responses. Nat Immunol 9, 733–742 (2008). |
| 8. | Charles, N., Hardwick, D., Daugas, E., Illei, G. G. & Rivera, J. Basophils and the T helper 2 environment can promote the development of lupus nephritis. Nat Med 16, 701–707 (2010). |
| 9. | Iikura, M. et al. Chemokine receptors in human basophils: inducible expression of functional CXCR4. J Leukoc Biol 70, 113–120 (2001). |
| 10. | Bischoff, S. C. Monocyte chemotactic protein 1 is a potent activator of human basophils. J Exp Med 175, 1271–1275 (1992). |
| 11. | Hirai, H. et al. Prostaglandin D2 selectively induces chemotaxis in T helper type 2 cells, eosinophils, and basophils via seven-transmembrane receptor CRTH2. J Exp Med 193, 255–261 (2001). |
| 12. | Satoh, T. et al. Prostaglandin D2 plays an essential role in chronic allergic inflammation of the skin via CRTH2 receptor. J Immunol 177, 2621–2629 (2006). |
| 13. | de Paulis, A. et al. Urokinase induces basophil chemotaxis through a urokinase receptor epitope that is an endogenous ligand for formyl peptide receptor-like 1 and -like 2. J Immunol 173, 5739–5748 (2004). |
| 14. | Lim, L. H. K. et al. Stimulation of human endothelium with IL-3 induces selective basophil accumulation in vitro. J Immunol 176, 5346–5353 (2006). |
| 15. | Kim, S. et al. Cutting edge: basophils are transiently recruited into the draining lymph nodes during helminth infection via IL-3, but infection-induced Th2 immunity can develop without basophil lymph node recruitment or IL-3. The Journal of Immunology 184, 1143–1147 (2010). |
| 16. | Siracusa, M. C. et al. TSLP promotes interleukin-3-independent basophil haematopoiesis and type 2 inflammation. Nature 477, 299–234 (2011). |
| 17. | Kim, S., Shen, T. & Min, B. Basophils can directly present or cross-present antigen to CD8 lymphocytes and alter CD8 T cell differentiation into IL-10-producing phenotypes. The Journal of Immunology 183, 3033–3039 (2009). |
| 18. | Pecaric-Petkovic, T., Didichenko, S. A., Kaempfer, S., Spiegl, N. & Dahinden, C. A. Human basophils and eosinophils are the direct target leukocytes of the novel IL-1 family member IL-33. Blood 113, 1526–1534 (2009). |
| 19. | Komai-Koma, M. et al. Interleukin-33 amplifies IgE synthesis and triggers mast cell degranulation via interleukin-4 in naïve mice. Allergy 67, 1118–1126 (2012). |
| 20. | Miura, K. & MacGlashan, D. W. Dual phase priming by IL-3 for leukotriene C4 generation in human basophils: difference in characteristics between acute and late priming effects. J Immunol 164, 3026–3034 (2000). |
| 21. | Didichenko, S. A., Spiegl, N., Brunner, T. & Dahinden, C. A. IL-3 induces a Pim1-dependent antiapoptotic pathway in primary human basophils. Blood 112, 3949–3958 (2008). |
| 22. | Ohmori, K. et al. IL-3 Induces Basophil Expansion In Vivo by Directing Granulocyte-Monocyte Progenitors to Differentiate into Basophil Lineage-Restricted Progenitors in the Bone Marrow and by Increasing the Number of Basophil/Mast Cell Progenitors in the Spleen. The Journal of Immunology 182, 2835–2841 (2009). |
| 23. | Spiegl, N., Didichenko, S., McCaffery, P., Langen, H. & Dahinden, C. A. Human basophils activated by mast cell-derived IL-3 express retinaldehyde dehydrogenase-II and produce the immunoregulatory mediator retinoic acid. Blood 112, 3762–3771 (2008). |
| 24. | Tschopp, C. M. et al. Granzyme B, a novel mediator of allergic inflammation: its induction and release in blood basophils and human asthma. Blood 108, 2290–2299 (2006). |
| 25. | Schroeder, J. T., Chichester, K. L. & Bieneman, A. P. Human basophils secrete IL-3: evidence of autocrine priming for phenotypic and functional responses in allergic disease. The Journal of Immunology 182, 2432–2438 (2009). |