Tetraspanins are modulators of mast cell degranulation
Mast cell (MC) degranulation basically is the result of FceRI
cross-linking by IgE bound antigen. However, this process is mediated by
a complex signaling machinery, sometimes referred to as the FceRI
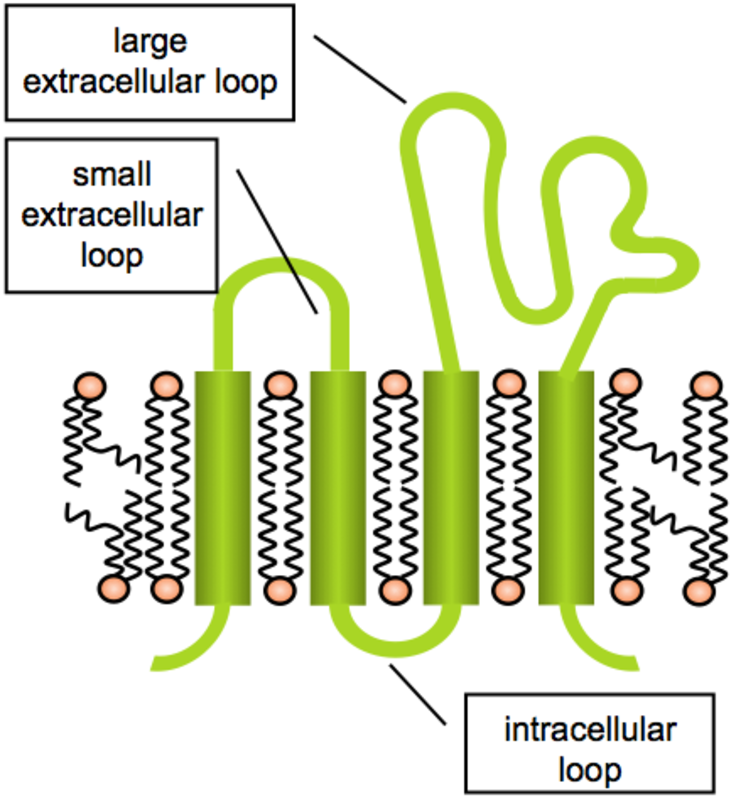
signalosome (1). Tetraspanin proteins have been named after their four
transmembrane domains that incorporate several conserved motifs (2).
They are often described as scaffold proteins that enable clustering of
other membrane proteins, such bringing together various partners of
intracellular signaling cascades (3) or increasing the density of some
receptors (e.g. integrins) at some location at the extracellular side of
the membrane (4). But tetraspanins are not restricted to the plasma
membrane, they also reside in the membranes of intracellular
compartments. An example for this is CD63, that is residing in the
membranes of granules of various cell types as well and has long been
known as an activation marker of human mast cells and basophils (5).
However, CD63 and other tetraspanins are not just markers, but also
modulators of MC degranulation by mediating clustering of FceRI and
co-clustering of additional membrane molecules important for successful
FceRI signaling. Surprisingly, they fine tune also additional MC
functions (6).
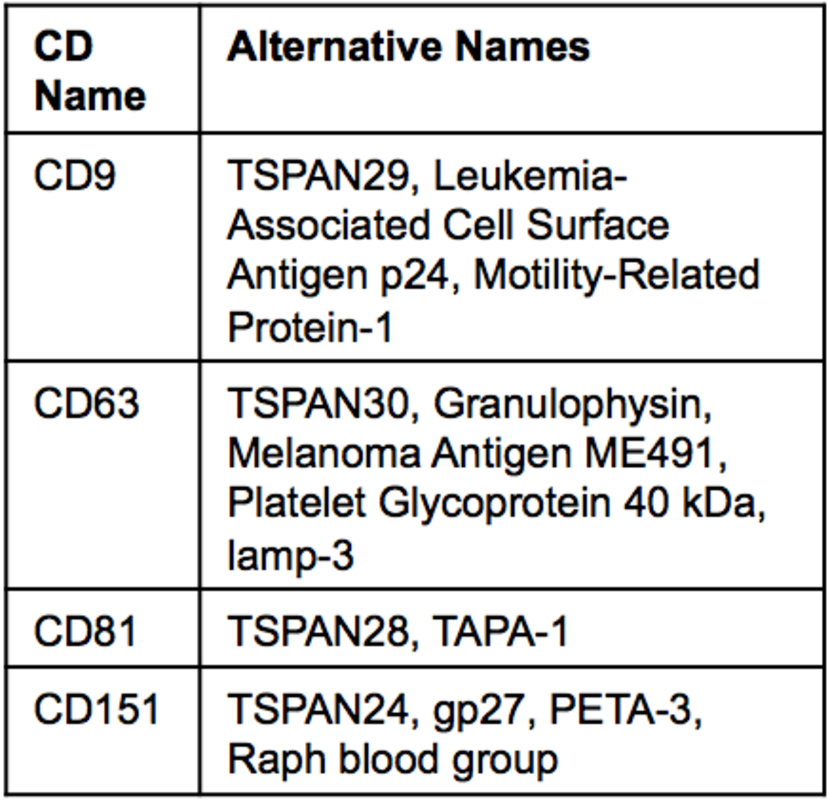
CD63
After FceRI crosslinking by IgE-Antibody complexes, CD63 containing
membranes of MC or basophil granules fuse with the plasma membrane in
the process of granule exocytosis. Interestingly, CD63 of granule origin
possesses a glucosylation pattern different from CD63 molecules that
always have resided in the plasma membrane. A new antibody specifically
recognizes this CD63 of granular origin. It has been used to demonstrate
that MC are able to degranulate more often than just once (7). CD63 is
supposed to be co-localized with the FceRI. A blocking antibody against
CD63 inhibits MC adhesion to the extracellular matrix (ECM) proteins
fibronectin and vitronectin, possibly by disrupting integrin-clustering
by CD63. It also neutralizes the degranulation promoting effect of MC
adhesion to ECM, supposedly by inhibition of the Gab2-PI3K pathway that
is a mediator of both adhesion and degranulation (8).
CD9
CD9 has been found to be co-localized with FceRI, but there is only
artificial evidence of modulation of degranulation by CD9 in RBL cells
(9). Instead, CD9 has been demonstrated to act as the main interleukin
16 (IL-16) receptor of MC and to mediate MC chemotaxis towards an IL-16
gradient (10).
CD81
A monoclonal antibody recognizing CD81 has been shown to inhibit mast cell degranulation after FceRI stimulation. As has been the case with the CD63 blocking antibody, the inhibitory effect was not mediated by inhibition of FceRI tyrosin phosphorylation, calcium influx, or leukotriene synthesis. Notably, the use of this antibody inhibited passive cutaneous anaphylaxis in a rat model (11).
CD151
The Tetraspanin CD151 seems to be expressed on most types of human mast cells and basophils, however there are no data showing a functional role of CD151 in these cells yet (6).
References:
| 1. | Draber P, Halova I, Levi-Schaffer F, Draberova L (2012) Front Immun 2:95. |
| 2. | Hemler ME (2005) Nat Rev Mol Cell Biol 6, 801-811. |
| 3. | Levy S, Shoham T (2005) Nature reviews Immunology 5, 136-148. |
| 4. | Berditchevski F (2001) J Cell Sci 114, 4143-4151. |
| 5. | Hennersdorf F, Florian S, Jakob A, Baumgartner K, Sonneck K, et al. (2005) Cell Res 15, 325-335. |
| 6. | Koberle M, Kaesler S, Kempf W, Wolbing F, Biedermann T (2012) Frontiers in immunology 3, 106. |
| 7. | Schafer T, Starkl P, Allard C, Wolf RM, Schweighoffer T (2010) Allergy 65, 1242-1255. |
| 8. | Kraft S, Fleming T, Billingsley JM, Lin SY, Jouvin MH, et al. (2005) J Exp Med 201, 385-396. |
| 9. | Higginbottom A, Wilkinson I, McCullough B, Lanza F, Azorsa DO, et al. (2000) Immunology 99, 546-552. |
| 10. | Qi JC, Wang J, Mandadi S, Tanaka K, Roufogalis BD, et al. (2006) Blood 107, 135-142. |
| 11. | Fleming TJ, Donnadieu E, Song CH, Laethem FV, Galli SJ, et al. (1997) J Exp Med 186, 1307-1314. |


{kind=link}
{kind=link}