I. Introduction
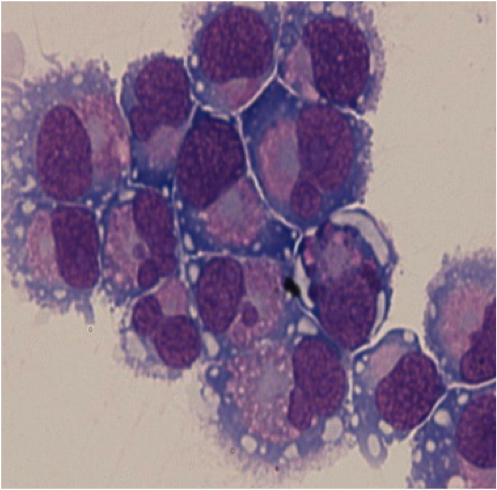
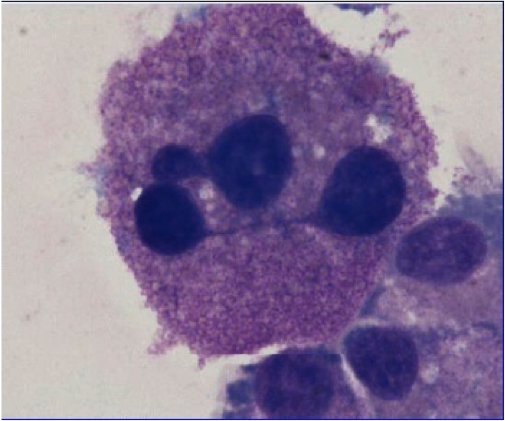
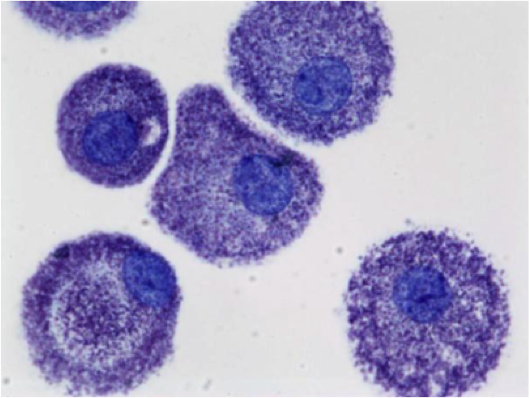
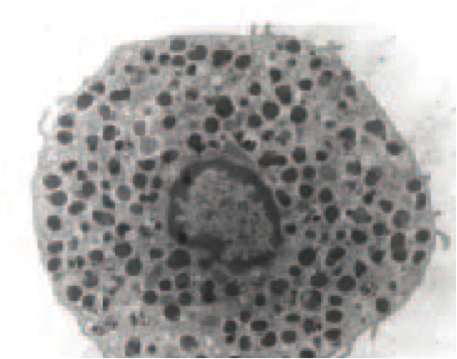
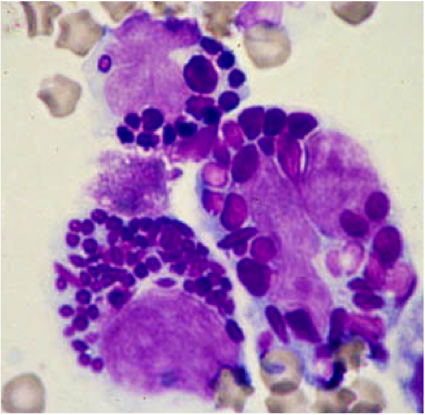
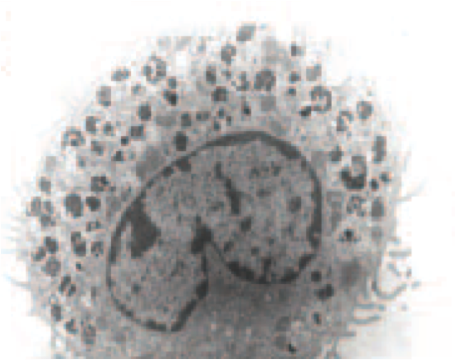
Mast cell (MC) progenitors, derived from hematopoietic progenitors (HP), circulate in the bloodstream and reach tissues to undergo terminal maturation upon the influence of cytokines, the most important being stem cell factor (SCF), whose receptor, KIT, is a transmembrane protein with tyrosine kinase (TK) activity. MCs are heterogeneous in morphology and functions between species but also between tissues in the same animal. In rodent, in mucosal tissues, MCs are more immature than in serosal tissues. In human, such heterogeneity is also found. Indeed, in mucosa, human MCs express only tryptase (MCT), whereas in connective tissues, they express tryptase and chymase (MCTC). MCs are pivotal players in innate immunity since they contribute to anti-infectious defense by direct interaction with pathogens or indirectly through ligation of bacterial compounds on Toll-like receptors (TLRs). They are involved in the physiopathology of allergic (through their high-affinity IgE receptors (FceRI)) and in auto-immune diseases (through their IgG receptors). Finally, MCs are involved in mastocytosis, a heterogeneous group of diseases characterized by their abnormal accumulation in tissues. In most cases, MCs found in mastocytosis express KIT defects.
Although widely distributed in the body, MCs represent a minor cell population. Methods based on differential sedimentation in high-density medium followed by purification by immunoaffinity allow to obtain relatively pure ex vivo mature MCs, but with a poor yield and sometimes altered properties. For these reasons, transformed MC lines of various origins have been extensively used. A few mouse and human tumor MC lines have been described, often lacking FcεRI, but they are not suitable for addressing physiologic issues. Thus, cultured MCs have been derived from various sources of mouse, rat or human HP and have been used to analyze various aspects of the biology of these elements.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
II. MCs in primary cultures
1. In rodents
Mouse and rat Bone Marrow-derived MCs (mBMMC and rBMMC)
Mouse BMMCs is the most widely used model. Mouse bone marrow (BM) cells cultured in the presence of recombinant murine (rm) IL-3 give rise, after 3 weeks, to virtually pure (>98%) Fcsym
RI+, Kit+ MCs that can keep growing for at least 2-3 months. Addition of recombinant murine SCF (rmSCF) to culture skews the phenotype from largely mucosal-like (IL-3) to more serosal-like (SCF). These mBMMCs have many advantages since they are easy to grow in large quantities and to maintain in culture costly. Techniques are available to transfect these cells and, finally, they can be used to replenish MC populations in MC-deprived animals.
Rat BMMCs are obtained by culture of rat BM cells in medium containing rrIL-3, but they do not proliferate as much and the culture, made by nearly pure immature MCs, declines around the fifth week. The rBMMCs obtained exhibit a limited number of large metachromatic granules, which can be released after FcεRI aggregation.
Mouse Peritoneal Cell-derived MCs (mPCMC)
One limitation of mBMMCs is their “mucosal-like” phenotype. To overcome this limitation, a technique has been developed where peritoneal cells are cultured in the presence of rmSCF. At two weeks and later, cultures consist of virtually pure (>98%) FcεRI+, Kitlow MCs. MPCMC thus obtained proliferate for about one month then stop growing. They keep alive for another 2-3 months, provided that they are maintained in rmSCF-containing medium. These cells are mature serosal-type mouse MCs and respond to IgG immune complexes through their FcγRIIIA. They are negatively regulated by FcγRIIB. PCMC also respond to IgE Abs, but IgE-induced mPCMC responses differ quantitatively and qualitatively from BMMC responses.
2. In human
Although some cell culture systems allow, after purification from tissues, the maintenance and even some limited growth of human MCs, most of the works on the biology of these cells are done on in vitro derived MCs, by culture of normal hHP (CD34+ or CD133+ cells), purified mainly from cord blood (CB) or peripheral blood (PB). CD34+ or CD133+ cells cultured in medium containing rhSCF and rhIL-6 (+rhIL-3 added first week only) initially proliferate 10 to 100 times or more during the first 2-3 weeks, depending on the origin of the CD34+ cells used. After that cells are reefed once a week in fresh medium. Usually, 106 CD34+ cells from CB may give rise to 5-15 x107 CBMC at 8-10 weeks, whereas 106 CD34+ cells from PB give rise to 1-2 x107 PBDMC at 7 weeks, in both cases with a purity > 95%.
PB-derived human MCs (PBMCs) are relatively mature with a condensed non-segmented nucleus and numerous granules. They express spontaneously FcεRI and are tryptase+ chymase+. By contrast, CB-derived MCs (CBMCs) are relatively immature with a segmented nucleus and less abundant granules. They are tryptase+ chymase- and they do not express spontaneously significant amount of FcεRI, but incubation for 5 days with IgE and rhIL-4 induce their expression of FcεRI. PBMCs or CBMCs expressing FcεRI can degranulate and release cytokines upon aggregation of the receptor. They express a number of TLRs that can differentially activate them upon stimulation with their respective ligands.
III. MC lines
1. Rodent MC lines
There are currently 3 factor-independent rodent MC lines (mouse P815 and FMA-3, and rat basophilic leukemia (RBL)) and one IL-3 dependent mouse MC line (MC/9). Interestingly, the factor-independent cell lines harbor Kit defects conferring spontaneous activation of the receptor and lack the FcεRI, by contrast to MC/9 cells.
The most popular is RBL-1 that was chemically induced in rats. Although it expressed FceRI and displayed MC characteristics, it was not secretory. However, in 1976, a histamine secreting RBL-2H3 clone was isolated. Since then, the RBL-2H3 cells have been used extensively for the study of FcεRI interactions with its ligand and subsequent degranulation. They are also employed for assessment studies for MC stabilizers.
2. Human MC lines
Three human MC lines are currently available HMC-1, LAD, and LUVA:
HMC-1 is a factor-independent, fast growing, immature MC line derived from the PB of a patient suffering from MC leukemia. In fact, two subclones, HMC-1.1 and HMC-1.2, have been characterized. HMC-1.1 has a KIT V560G mutation, whereas HMC-1.2 has two KIT mutations (V560G and D816V). HMC-1 cells present with many other molecular abnormalities. The major limitations of HMC-1 are the absence of expression of the FcεRI and their paucity in granules.
LAD1 and LAD2 FcεRI+ cell lines were derived from the BM of a patient with MC sarcoma with KIT D816V mutation. Contrasting to the tumor cells of the patient, they have a normal KIT and remain dependent upon SCF for survival and proliferation. They express intracytoplasmic histamine, tryptase and chymase. LAD cells release b-hexosaminidase following FceRI or FcgRI aggregation, making it potentially interesting. However their variable but long doubling time (about 15 days) makes them poorly suitable for screening assays requiring large number of cells.
LUVA cell line was derived in 2011 from purified PB CD34+ cells of a donor with aspirin-exacerbated respiratory disease. They are granulated, tryptase+, but are growth factor independent, although having a normal KIT. They were initially FcεRI+ and able to release β-hexosaminidase, leukotrienes and cytokines. However, it seems that the cell line tends to lose the expression of FceRI upon long-term culture.
IV. Conclusion
Today, the cellular models representing human MCs have many limitations, either because they are poorly reproducible and expensive to produce (primary culture models) or because they are far away from MCs found in vivo in terms of biological properties (cell lines). Thus there is still a need to develop a relevant, reproducible and inexpensive in vitro model nearing at the best the main morphological and biological properties of MCs, allowing a better understanding of their functions in health and diseases.