Basophils and mast cells belong to distinct myeloid lineages, but they share a number of morphological and functional features (Stone 2010). Most notably, they are the only cell types which respond to immunoglobulin E (IgE)-induced activation via the high affinity receptor for the constant fragment epsilon (RFceI). IgE-dependent activation of mast cells and basophils leads to rapid and massive exocytosis of secretory granules contents, followed by lipid mediator generation and delayed cytokine responses. Mast cells and basophils also share a number of IgE-independent pathways of activation, for example through bacterial compounds such as formylated peptides, or pathogen-associated molecular patterns (PAMPs). Because basophils are readily identified among circulating leukocytes and on the basis of common activation mechanisms, basophils were assumed to be a convenient surrogate for the assessment of allergen-mast cell interaction in humans (Shelley 1963). Basophil activation can be monitored through morphological changes, phenotypic changes, secreted mediators quantification, or a combination of these (Ebo 2012).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CLINICAL IMPLICATIONS
Currently, basophil activation tests may contribute to (McGowan 2012): (i) diagnosis of IgE-dependent allergic responses to drugs or foods when no skin prick test (SPT) or serum IgE detection is available, or in cases of evocative clinical history with negative SPT and serum IgE; (ii) venom immunotherapy initiation and follow-up; (iii) preliminary in vitro testing in cases of severe systemic anaphylaxis, before challenging the patient with the culprit allergen; (iv) search for an autoimmune anti-IgE or anti-IgE receptor basophil activation in chronic urticaria assessment (Konstantinou 2012).
MICROSCOPY
The first basophil activation tests were made available as soon as 1962. By that time, immunoglobulin E had not been identified yet, but the first human basophil degranulation test intended for the diagnosis of allergy and anaphylaxis was reported by WB Shelley and L Juhlin (Shelley’s test, Shelley 1962). The first generation of basophil activation tests relied on degranulation monitoring by means of optical microscopy. Basophil granules are stained with basic dyes, such as toluidin blue, Alcian blue, or methylene blue, which bind granular proteoglycans. Degranulation following activation leads to the partial loss of granular staining. As a rule, 50 to 100 basophils were counted in control samples and after allergen challenges. Commercial methods using the basophil degranulation test as an allergy marker were available until the late 80s. The advantage of such methods is the specific granule staining for either basophils or mast cells; their major drawback in clinical settings is the limited number of basophils that one can count on peripheral blood smears. Moreover, microscopy-based tests are highly subjective. Therefore, basophil staining and visualisation rapidly took advantage of the 1990s major progress in flow cytometry techniques.
CYTOMETRY
Basophil counting and activation monitoring by means of flow cytometry became available in the 1990s (Knol 1991). Improved techniques are now available, providing a large choice for basophil workers in the medical and research fields. There are three major issues with cytometric basophil activation tests. First, basophils must be identified among other leukocytes, then, after applying a stimulus, activated and non activated basophils must be distinguished. The third issue is the need for high numbers of basophils to be analyzed, usually not less than 500-1000 cells.
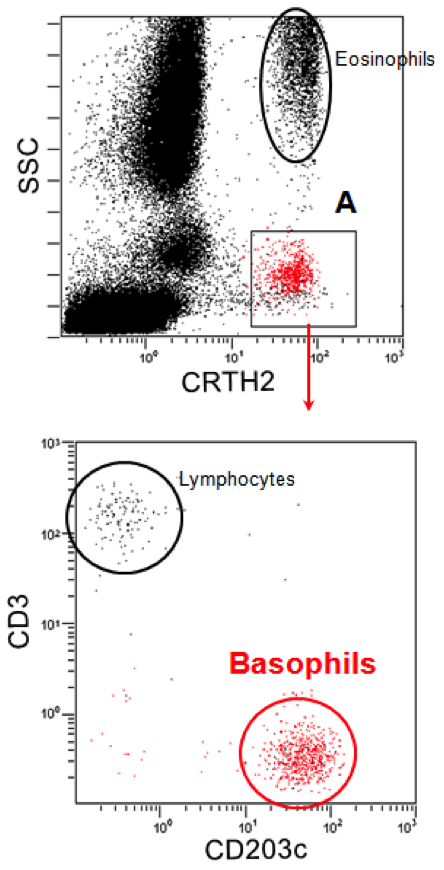
Basophil identification often relies on CCR3 (CD193, eotaxin-1 receptor) detection on plasma membrane, in combination with low side scattering of the light. Thus, small, non granular cells which are grossly located between the lymphocyte and the monocyte gates and express high levels of CCR3 are recognized as basophils. Large, granular CCR3+ cells mainly comprise eosinophils.
Detection of membrane-bound IgE on low side scatter cells was one of the earliest means of basophil identification among leukocytes. Taken alone, IgE detection may lack specificity, because of various IgE-bearing cells, such as monocytes in atopic patients, or eosinophils and dendritic cells. Moreover, IgE density on plasma membrane is highly heterogeneous among individuals.
A combination of IgE and CD203c detection on plasma membrane may be used for basophil identification. CD203c is expressed at low levels by resting basophils, and is rapidly upregulated at the plasma membrane of activated ones. Therefore, CD203c is a versatile marker for both basophil identification and activation monitoring.
Alternative strategies for basophil identification include combined CRTH2 (CD294, prostaglandin receptor) and CD3 staining and low side scattering. CRTH2 is expressed by basophils, eosinophils and Th2-type lymphocytes. Low side scattering excludes eosinophils, and positive CD3 further excludes lymphocytes.
Finally, basophils may be identified through CD123 (IL-3 receptor) positive staining and the lack of MHC class II staining.
For basophil activation monitoring, current methods usually rely on CD203c, CD63, or both. As previously noted, CD203c is a plasma membrane molecule constitutively expressed at low levels by resting basophils. CD203c is an ectonucleotide pyrophosphatase phospho-diesterase (E-NPP3) whose physiological relevance in basophils has remained as yet unclear. Plasma membrane CD203c undergoes very rapid upregulation after basophil activation (5-15 minutes) and mirrors early activation processes which may not involve complete basophil degranulation.
In contrast, CD63 is a tetraspanin molecule present in the lysosomal and secretory granule membranes. CD63 is not expressed at the plasma membrane of resting basophils , but reaches the plasma membrane as a result of granule translocation and membrane fusion. CD63 expression by degranulating basophils is slightly delayed compared to CD203c upregulation (10-25 minutes). IL-3 priming enhances basophil degranulation and CD63 expression.
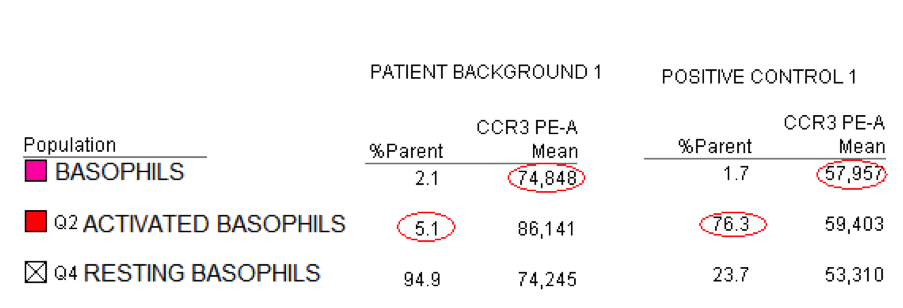
Finally, the CCR3 apparent drop in mean fluorescence intensity (MFI) during basophil activation may help with activation monitoring. CCR3 molecules are dispersed all around in the plasma membrane of resting basophils; they aggregate in lipid rafts during basophil activation, a phenomenon that leads to a diminished fluorescence level of CCR3 (Sainte-Laudy 2010). The accuracy of basophil identification and activation monitoring through flow cytometric means allowed cytometry tests to enter routine laboratory work-up of allergic patients.
SECRETED MEDIATORS
Histamine release, which is responsible in vivo for vasodilation, brochoconstriction, and itching, was first monitored as a product from mast cells and basophils, either in vivo or in vitro, in the early 1960s (Schauer&Werle, 1959; Juhlin&Shelley, 1966). A popular test was the radio-immune assay of histamine in biological fluids based on binding competition between unlabeled histamine in the biological sample and 125I histamine to an anti-histamine antibody coated to the test plate or tube (Broide 1990). Although highly sensitive, histamine release tests for the diagnosis of systemic anaphylaxis lack specificity . Pregnancy, hemolysis, or inadequate tube storage and transport are among the most common causes of false positive results, while delayed blood collection is the major cause of false negative results. Tryptase, a mast cell specific enzyme, has almost completely replaced histamine in the laboratory work-up of systemic anaphylaxis (Brown 2011). Currently, most histamine assays are performed with ELISA methods.
Another preformed mediator of basophils and mast cells is beta-hexosaminidase, an acid hydrolase that characterizes lysosomal-derived secretory granules (Schwartz 1979). The beta-hexosaminidase release test is mainly used for research purposes with basophilic or mast cell lines.
For medical purposes, de novo synthesis of leukotrienes and prostaglandins can also be used to monitor basophil activation. Basophils produce large amounts of leukotriene C4 (LTC4), which can be measured in supernatants of basophils following a 20-minute stimulation (MacGlashan 1986).
COMBINED METHODS
Basophil activation may be assessed through combined strategies, thus increasing the sensitivity and the specificity of the tests. Phenotypic markers of activation may be coupled to leukotriene assay in the supernatant; alternatively, histamine release and plasma membrane expression of activation markers can be assayed simultaneously (Ebo 2012).
CONCLUDING REMARKS
Basophil activation was the first allergy test to be published 50 years ago. The complexity of basophil pathophysiology and the technical variability among laboratories explain most of the as-yet unresolved issues of its standarization. Yet, basophil activation tests may help in allergy and anaphylaxis management, and may become a reliable surrogatefor allergen challenge tests in the future.
REFERENCES
- Broide DH, Smith CM, Wasserman SI. Mast cells and pulmonary fibrosis. Identification of a histamine releasing factor in bronchoalveolar lavage fluid. J Immunol 1990, 145:1838-44.
- Brown SGA, Stone SF. Laboratory diagnosis of acute anaphylaxis. Clin Exp Allergy 2011,41:1660-2.
- Ebo DG, Bridts CH, Mertens CH et al. Analyzing histamine release by flow cytometry (HistaFlow): a novel instrument to study the degranulation pattern of basophils. J Immunol Methods 2012, 375:30-8.
- Knol EF, Mul FP, Jansen H et al. Monitoring human basophil activation via CD63 monoclonal antibody 435. J Allergy Clin Immunol 1991, 88:328-38.
- Konstantinou GN, Asero R, Ferrer M et al. EAACI taskforce position paper: evidence for autoimmune urticaria and proposal for defining diagnostic criteria. Allergy 2012, online.
- MacGlashan DW, Peters SP, Warner J et al. Characteristics of human basophil sulfidopeptide leukotriene release: releasability defined as the ability of the basophil to respond to dimeric cross-links. J Immunol 1986,136:2231-9.
- McGowan EC, Saini S. Update on the performance and application of the basophil activation tests. Curr Allergy Asthma Rep 2012, online.
- Sainte-Laudy J, Ouk C. Use of lipid rafting for the analysis of human basophil activation by flow cytometry. Infl Res 2010,59:S193-5.
- Schauer A, Werle E. Zur histochemische Darstellung des Histamins der Mastzellen. Z Ges Exp Med 1959, 131:100-4.
- Schwartz LB, Austen KF, Wasserman SI. Immunologic release of beta-hexosaminidase and beta-glucuronidase from purified rat serosal mast cells. J Immunol 1979,123:1445-50.
- Shelley WB, Caro WA. Detection of anaphylactic sensitivity by the basophil degranulation response. An experimental study. JAMA, 1962 ; 182 : 172-8
- Shelley WB, Juhlin L. Functional cytology of the human basophil in allergic and physiologic reactions: technic and atlas. Blood 1962, 19:208-16.
- Shelley WB. Methods of observing the basophil leukocyte degranulation response. Ann NY Acad Sci 1963,103:427-35.
- Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol 2010,125:S73-80.