Mast cells are emerging players in physiological and pathological pain pathways. Mast cells are frequently found in close proximity to nociceptive neurons and therefore can participate in juxtacrine signaling in neuro-immune synapses (1). Mast cells induce nociceptor activation through the release of chemical mediators during degranulation and can be activated by mediators released from nociceptors upon injury (2).
In a model of passive cutaneous anaphylaxis (PCA) in the rat hind paw Lavich and colleagues (3) demonstrated transient increased thermal hyperalgesia following plantar anti-DNP-IgE sensitization and DNP challenge regulated by the synergistic interaction of histamine, bradykinin, and 5-hydroxytryptamine – all likely products of mast cell degranulation (3). Piovezan and colleagues observed abrogation of OVA-induced hyperalgesia by chemical depletion of mast cells in the hind paws of sensitized mice (4), demonstrating that active sensitization and challenge leads to pain that is dependent on mast cell degranulation. The injection of IL-6, TNFα, IL-1β, and IL-8, cytokines readily secreted by mast cells, can independently evoke hyperalgesia (5). Chatterjea and colleagues showed that genetically mast cell-deficient C57BL/6-KitW-sh/W-sh mice have significantly reduced thermal and mechanical plantar hyperalgesia after intra-plantar injection with mast cell secretagogue compound 48/80 (c48/80) compared to wild-type mice; these responses are restored following reconstitution of the hind paw with bone marrow cultured mast cells (6). These findings supported an earlier observation that injection of c48/80 into the forearms of young male volunteers caused changes in pain sensitivity in the skin surrounding the sites of administration (7).
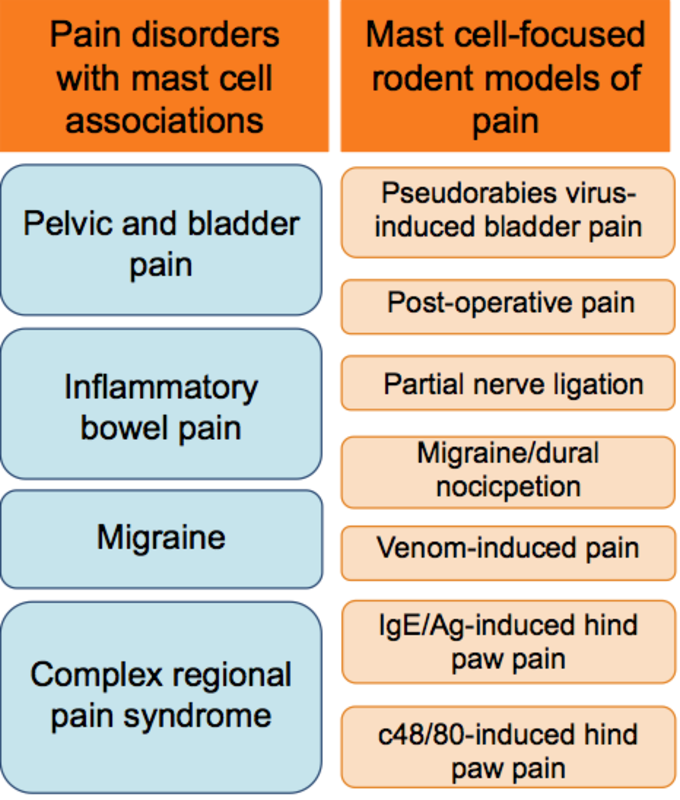
Mast cells in pre-clinical models of inflammatory and chronic pain disorder:
Mast cells have been shown to be important in rodent models of migraine (16, 17) with systemic mast cell degranulation causing nociceptor activation in the dura as well as in the spinal cord. Mast cells contribute to mechanical cystitis pain via histamine receptor 1 and 2 signaling in a murine model of pseudorabies virus (PRV)-induced pelvic pain (18). NK-1 and histamine receptor 2 antagonists have been shown to mitigate bladder-associated pelvic pain, suggesting that blockade of mast cell-mediated regulatory mechanisms may have therapeutic potential in the treatment of cystitis-related pain (19). Bonavita and colleagues demonstrated that mast cell stabilizer sodium cromoglycate (SCG) inhibited Bothrops jararaca venom-induced release of histamine and leukotriene C4 in vitro and abrogated venom-induced hyperalgesia in vivo in rats (20). Mast cell are also critical for pain sensation in rodent models of post-operative pain (21) and neuropathic thermal and mechanical hyperalgesia caused by the ligation of the sciatic nerve in rats (22).
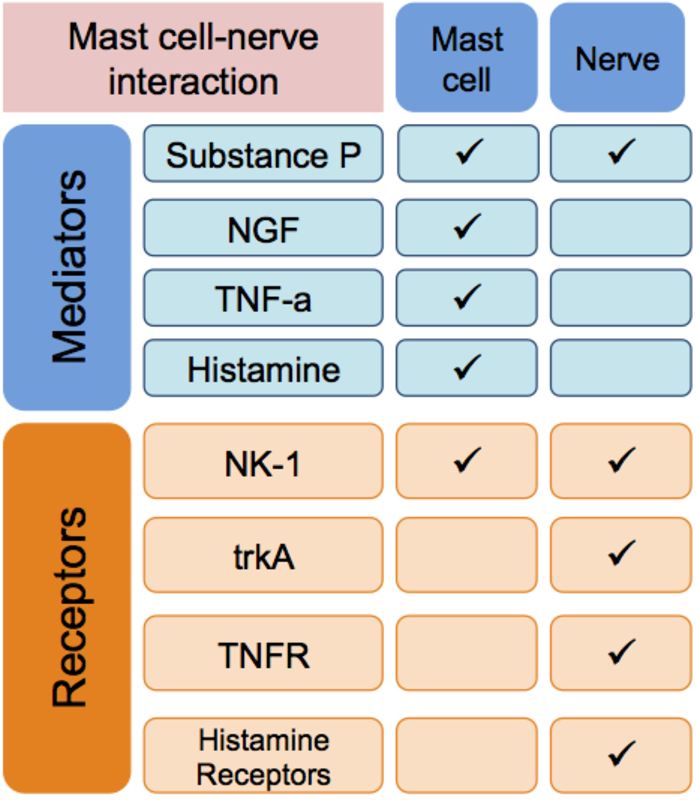
Mast cell-neuron interactions in pain pathways:
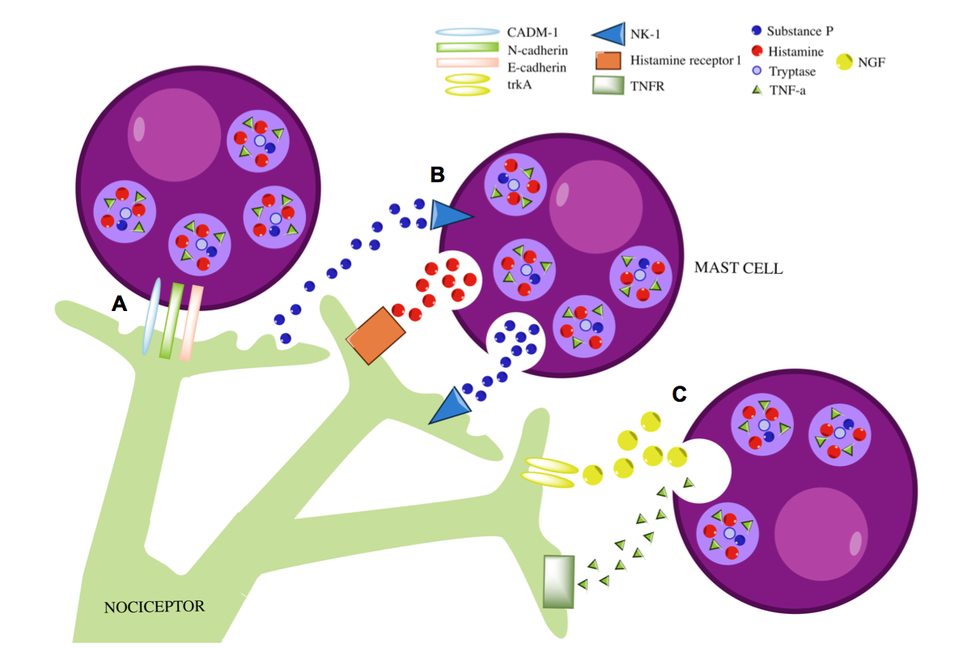
Mast cells residing in close proximity to unmyelinated nerve fibers, such as the nociceptive C-fibers, can undergo ultrastructural alterations that allow differential or selective “piecemeal degranulation” (3). This is particularly important for understanding pain conditions where mast cell-nerve associations have been documented, such as inflammatory bowel syndrome (16) and vulvodynia (12, 14). Adhesion molecules such as N-cadherin and cell adhesion molecule-1 (CADM-1) facilitate mast cell-nerve junctions (23). Mast cell mediators NGF and TNF-a influence neuronal growth, while mast cell-derived NGF has also been shown to lower the threshold of nociceptor firing through its binding to TRK1-transforming tyrosine kinase protein (trkA) (24, 25). Another important signal at the mast cell-nerve synapse is substance P. Released by both neurons and mast cells, substance P leads to production of prostaglandins and leukotrienes, as well as TNF-a and IL-6 by mast cells (26, 27). Substance P-NK-1 binding primes mast cells to degranulate upon repeated application of lower doses of substance P (28).
Conclusion:
Mast cells reside in sentinel locations in the tissue and release a versatile repertoire of mediators (29) including those that interact structurally and functionally with pain-sensing nociceptors (3). Their contributions to pain pathways in the peripheral tissues as well as in central sensitization mechanisms are active areas of research. These investigations will contribute to our improved understanding of mast cell biology and to the design of novel, rational therapies for the treatment and management of pain.
Acknowledgements:
Tijana Martinov and Alyssa Ashbaugh contributed to this entry.
References:
| 1. | Ren, K., R. Dubner. 2010. Interactions between the immune and nervous systems in pain. Nat Med. 16(11): 1267-76. |
| 2. | Forsythe, P., J. Bienenstock. 2012. The mast cell-nerve functional unit: a key component of physiologic and pathophysiologic responses. Chem Imunol Allergy. 98(1): 196-221. |
| 3. | Lavich, T. R., R. S.Cordeiro, J. B.Calixto, P. M. Silva, M. A. Martins. 2003. Combined action of vasoactive amines and bradykinin mediates allergen-evoked thermal hyperalgesia in rats. Eur J Pharmacol. 462(1-3): 185-92. |
| 4. | Piovezan, A.P., P. D’orléans-Juste, M. Frighetto, G. E. Souza, M. G. Henriques, G. A. Rae. 2004. Endothelins contribute towards nociception induced by antigen in ovalbumin-sensitised mice. Br J Pharmacol. 141(4): 755-63. |
| 5. | Verri, W., T. Cunha, C. Parada, S. Poole, F. Cunha, S. Ferreira. 2006. Hypernociceptive role of cytokines and chemokines: targets for analgesic drug development? Pharmacology & Therapeutics. 112(1): 116-38. |
| 6. | Chatterjea, D., A. Wetzel, M. Mack, et al. 2012. Mast cell degranulation mediates compound 48/80-induced hyperalgesia in mice. Biochem Biophys Res Commun. 425(2): 237-43. |
| 7. | Drummond, P. D. 2004. The effect of cutaneous mast cell degranulation on sensitivity to heat. Inflamm Res. 53(7): 309-15. |
| 8. | Theoharides, T. C., J. Donelan, K. Kandere-Grzybowska, A. Konstantinidou. 2005. The role of mast cells in migraine pathophysiology. Brain Res Brain Res Rev. 49(1): 65-76. |
| 9. | Done, J.D., C.N. Rudick, M.L. Quick, A.J. Schaeffer, P. Thumbikat. 2012. Role of mast cells in male chronic pelvic pain. J Urol. 187(4): 1473-82. |
| 10. | Li, W., T. Z. Guo, D. Y. Liang, Y. Sun, W. S. Kingery, J. D. Clark. 2012. Substance P signaling controls mast cell activation, degranulation, and nociceptive sensitization in a rat fracture model of complex regional pain syndrome. Anesthesiology. 116(4): 882-95. |
| 11. | Bornstein, J., N. Goldschmid, E. Sabo. 2004. Hyperinnervation and mast cell activation may be used as histopathologic diagnostic criteria for vulvar vestibulitis. Gynecol Obstet Invest. 58(3): 171-8. |
| 12. | Bornstein, J., Y. Cohen , D. Zarfati, S. Sela, E. Ophir. 2008. Involvement of heparanase in the pathogenesis of localized vulvodynia. Int J Gynecol Pathol. 27(1): 136-41. |
| 13. | Symons, F. J., G. Wendelschafer-Crabb, W. Kennedy, W. Heeth , J. W. Bodfish. 2009. Degranulated mast cells in the skin of adults with self-injurious behavior and neurodevelopmental disorders. Brain Behav Immun. 23(3): 365-70. |
| 14. | Barbara, G., V Stanghellini, R. De Giorgio, R. Corinaldesi. 2006. Functional gastrointestinal disorders and mast cells: implications for therapy. Neurogastroenterol Motil. 18(1): 6-17. |
| 15. | Levy, D., R. Burstein, V. Kainz, M.Jakubowski, A.M. Strassman. 2007. Mast cell degranulation activates a pain pathway underlying migraine headache. Pain. 130(1-2): 166-76. |
| 16. | Levy, D., V. Kainz, R. Burstein, A. M. Strassman. 2011. Mast cell degranulation distinctly activates trigemino-cervical and lumbosacral pain pathways and elicits widespread tactile pain hypersensitivity. Brain Behav Immun. 26(2): 311-7. |
| 17. | Rudick, C. N., P. J. Bryce, L. A. Guichelaar, R. E. Berry, D. J. Klumpp. 2008. Mast cell-derived histamine mediates cystitis pain. PLOS ONE. 3(5): e2096. |
| 18. | Rudick, C.N., A. J. Schaeffer, D. J. Klumpp. 2009. Pharmacologic attenuation of pelvic pain in a murine model of interstitial cystitis. BMC Urol. 9(16): 1-8. |
| 19. | Bonavita, A. G., A. S. Da Costa, A. L. Pires, et al. 2006. Contribution of mast cells and snake venom metalloproteinases to the hyperalgesia induced by Bothrops jararaca venom in rats. Toxicon. 47(8): 885-93. |
| 20. | Zuo, Y., N. M. Perkins, D. J. Tracey, C. L. Geczy. 2003. Inflammation and hyperalgesia induced by nerve injury in the rat: a key role of mast cells. Pain. 105(3): 467-79. |
| 21. | Oliveira, S. M., C. C. Drewes, C. R. Silva, et al. 2011. Involvement of mast cells in a mouse model of postoperative pain. Eur J Pharmacol. 672(1-3): 88-95. |
| 22. | Suzuki, A., R. Suzuki, T. Furuno, R. Teshima, M. Nakanishi. 2004. N-Cadherin plays a role in the synapse-like structures between mast cells and neurites. Biol. Pharm. Bull. 27(12): 1891-4. |
| 23. | Leon, A., A. Buriani, R. Dal Toso, M. Fabris, S. Romanello, L. Aloe, R. Levi-Montalcini. 1994. Mast cells synthesize, store, and release nerve growth factor. Neurobiology. 91(1): 3739-43. |
| 24. | Kakurai, M., R. Monteforte, H. Suto, M. Tsai, S. Nakae, S. Galli. 2006. Mast cell-derived tumor necrosis factor can promote nerve fiber elongation in the skin during contact hypersensitivity in mice. The American Journal of Pathology. 169(5): 1713-21. |
| 25. | van Houwelingen, A.H., M. Kool, S.C.A. de Jager, F.A.M. Redegeld, D. van Heuven-Nolsen, A.D. Kraneveld, F.P. Nijkamp. 2002. Mast cell-derived TNF-TNF-α primes sensory nerve endings in a pulmonary hypersensitivity reaction. J Immunol. 168(1): 5297-302. |
| 26. | Kawabata A. 2011. Prostaglandin E2 and pain–an update. Biol Pharm Bull. 34(8):1170-3. Review. |
| 27. | Okuse K. 2007. Pain signalling pathways: from cytokines to ion channels. Int J Biochem Cell Biol. 39(3):490-6. Review. |
| 28. | Janiszewski J, Bienenstock J, Blennerhassett MG. 1994. Picomolar doses of substance P trigger electrical responses in mast cells without degranulation. Am J Physiol. 267:C138-45. |
| 29. | Galli SJ, Kalesnikoff J, Grimbaldeston MA, Piliponsky AM, Williams CM, Tsai M. 2005. Mast cells as “tunable” effector and immunoregulatory cells: recent advances. Annu Rev Immunol. 23:749-86. Review. |

{kind=link}
{kind=link}
{kind=link}