Mast cells receptors - general description
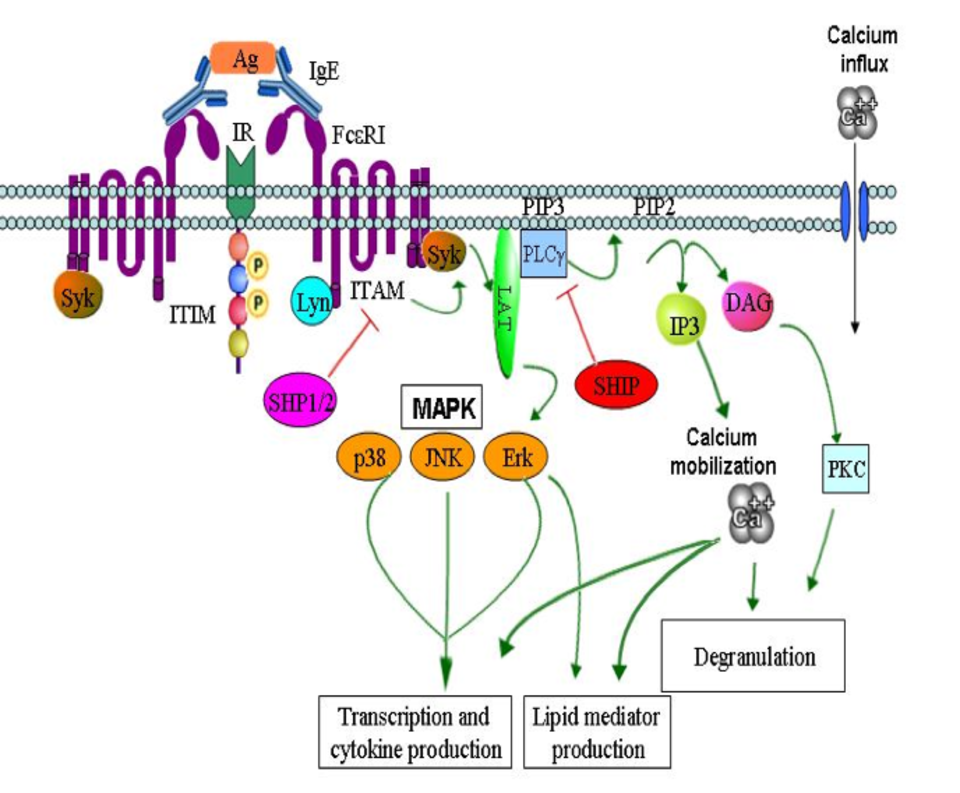
Mast cells (MC) are key effector cells in allergic inflammation (AI) but are also involved in regulating fibrosis, in tissue responses to neoplastic diseases, in autoimmune diseases and in host‑defense against bacterial and parasitic infections [1]. In allergy, the main mechanism of MC activation is through the binding of an allergen to the FcεRI‑IgE complex and transmission of signals via immunoreceptor tyrosine‑based activation motifs (ITAM). Activation signals are counterbalanced by negative or inhibition signals transmitted by immunoreceptor tyrosine‑based inhibition motifs (ITIM) bearing receptors; namely Inhibitory receptors (IRs). Classical ITIMs are defined as a consensus of a six amino acid sequence, I/V/LxYxxL/V (x denoting any amino acid), present in the intracytoplasmic domain of transmembrane molecules with inhibitory properties [2]. IRs belong either to the Ig receptor super family characterized by a single V-type Ig-like domain in the extracellular portion, or to the c-type (calcium-dependent) lectin super family. Upon activation, these receptors undergo tyrosine phosphorylation, often by a kinase of the Src family, which provides a docking site for the recruitment of cytoplasmic phosphatases having an SH2 domain such as SHP-1, SHP-2 and SHIP (figure 1; red arrow indicating the inhibitory pathway). IRs were found to be expressed on many cell types including NK cells, neutrophils, basophils, B cells and more. On MC, both murine bone marrow derived (BMMC) and human cord blood derived (CBMC), many IRs have been identified. These IRs include ones from the Ig-superfamily such as FCγRIIB, various siglecs, CD300a, SIRP-α, LILR(Gp49B1),PIR-B, LAIR-1, CD200R and PECAM-1. Other IRs belonging to c-type lectin including MAFA and CD72 [3] [4].
Inhibiting receptors, represented by CD300a receptor
The inhibitory receptor CD300a belongs to the Ig-superfamily and is one of seven members in a family of stimulatory as well as inhibitory type I transmembrane glycoproteins that control and modulate leukocyte responses. CD300a contains four ITIMs in its cytoplasmic tail which mediate its inhibitory function. While it was initially discovered on NK cells [5], CD300a expression and function has also been demonstrated in many cell types such as MC [6], T cell subsets, eosinophils [7], basophils [8] [9], monocytes and dendritic cells [10]. Importantly, CD300a inhibitory function was evident in human CBMC as well as in murine BMMC which express the murine orthologue namely Lmir-1. In CBMC, co‑aggregation of CD300a with IgE‑bound FcεRI, led to inhibition of IgE‑induced (but not of compound 48/80‑induced) b‑hexosaminidase, tryptase and IL-4 release. Additionally, CD300a aggregation led to the inhibition of CBMC Scf-mediated survival.
When using the bi-specific α‑Kit/α‑CD300a Ab, Kit‑mediated CBMC differentiation, survival and activation; and IgE dependent activation were abrogated. In the context of MC‑related disorders, when added to the malignant human MC line HMC‑1,
where Kit is constitutively activated, the CD300a bi-specific Ab inhibited mediator release without affecting cell survival [6].
In murine models of allergic diseases such as peritonitis, passive cutaneous anaphylaxis and acute asthma, the use of the murine bi‑specific Ab α‑IgE/ α-CD300a (Lmir‑1) proved to be effective in abrogating these allergic reactions [11]. Recently, a ligand for CD300a was identified as phosphatidylserine (PS), which is exposed on the outer leaflet of the plasma membrane of apoptotic cells, indicating CD300a as a new PS receptor [12].
Pharmacology
The expression of IRs on MC “makes them” attractive targets for new drug designs for allergy as well as other MC‑associated diseases such as mastocytosis (MC-associated cancers). Moreover, some of the studies on IRs, intended for therapeutic use nowadays are at an advanced stage. This is specifically important for the design of new and effective drugs to replace the current treatments which are mostly anti-symptomatic and lead to various unpleasant side effects.
References:
| 1. | Bachelet, I., F. Levi-Schaffer, and Y.A. Mekori, Mast cells: not only in allergy. Immunol Allergy Clin North Am, 2006. 26(3): p. 407-25. |
| 2. | Daeron, M., et al., Immunoreceptor tyrosine-based inhibition motifs: a quest in the past and future. Immunol Rev, 2008. 224: p. 11-43. |
| 3. | Karra, L., et al., Are we ready to downregulate mast cells? Curr Opin Immunol, 2009. 21(6): p. 708-14. |
| 4. | Karra, L. and F. Levi-Schaffer, Down-regulation of mast cell responses through ITIM containing inhibitory receptors. Adv Exp Med Biol, 2011. 716: p. 143-59. |
| 5. | Cantoni, C., et al., Molecular and functional characterization of IRp60, a member of the immunoglobulin superfamily that functions as an inhibitory receptor in human NK cells. Eur J Immunol, 1999. 29(10): p. 3148-59. |
| 6. | Bachelet, I., et al., The inhibitory receptor IRp60 (CD300a) is expressed and functional on human mast cells. J Immunol, 2005. 175(12): p. 7989-95. |
| 7. | Munitz, A., et al., The inhibitory receptor IRp60 (CD300a) suppresses the effects of IL-5, GM-CSF, and eotaxin on human peripheral blood eosinophils. Blood, 2006. 107(5): p. 1996-2003. |
| 8. | Sabato, V., et al., CD300a is expressed on human basophils and seems to inhibit IgE/FcepsilonRI-dependent anaphylactic degranulation. Cytometry B Clin Cytom, 2012. 82(3): p. 132-8. |
| 9. | Gibbs, B.F., Expressions and Inhibitory Functions of CD300a Receptors on Purified Human Basophils. Exp. Dermatol., 2012, [in press]. |
| 10. | Gasiorowski, R.E., et al., CD300 molecule regulation of human dendritic cell functions. Immunol Lett, 2012. |
| 11. | Munitz, A., I. Bachelet, and F. Levi-Schaffer, Reversal of airway inflammation and remodeling in asthma by a bispecific antibody fragment linking CCR3 to CD300a. J Allergy Clin Immunol, 2006. 118(5): p. 1082-9. |
| 12. | Nakahashi-Oda, C., et al., Identification of phosphatidylserine as a ligand for the CD300a immunoreceptor. Biochem Biophys Res Commun, 2012. 417(1): p. 646-50. |


{kind=link}