General Description
Besides the high affinity IgE receptor mast cells (MC) also express Fcγ receptors (FcγR) (1). However, other receptors for immunoglobulin isotypes such as FcaRI, FcdRI and Fca/mR are not found to be expressed on MC. FcγR exist in multiple isoforms categorized, either in terms of affinity as high-affinity and low-affinity receptors, or in terms of signalling pathways they induce, activating and inhibitory receptors. There are two high-affinity IgG receptors, FcγRI (CD64) and FcγRIV, and two families of low-affinity IgG receptors, FcγRII (CD32) and FcγRIII (CD16). FcγRII has 3 subtypes, i.e. FcγRIIA, FcγRIIB, and FcγRIIC. FcγRIII has 2 subtypes, i.e. FcγRIIIA and FcγRIIIB (2). The gene encoding human FcγRIIA generates two variants differing at position 131 (H131 and R131, with the former having a higher binding capacity for IgG) (3). FcγRIIB is described as an inhibitory receptor (3).
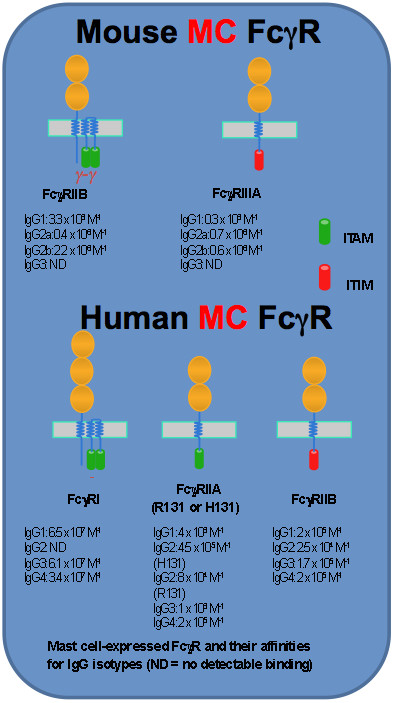
The expression patterns of FcγR and their structures and binding affinities
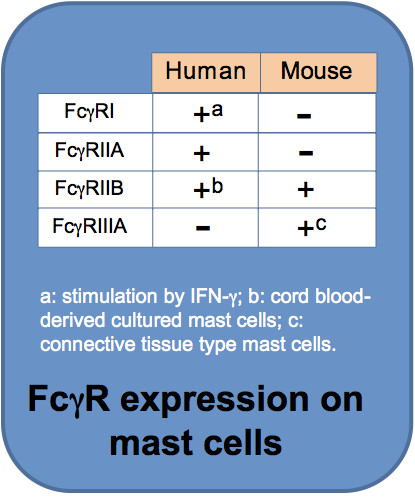
FcγR expression in MC is heterogenous. Discrepancies between species are well documented. FcγRI is found on cultured human MC upon interferon-g stimulation (4) or on MC from skin lesions in patients with psoriasis where interferon-g is enriched (5). Expression of FcγRIIA and FcγRIIB in human MC differs depending on tissue location (1). FcγRIIA, but not FcγRIIB, is expressed on human skin-derived MC (6). Human MC derived from umbilical cord blood express FcγRIIB (7). In mice, FcγR expression on MC is also subtype-specific. FcγRIIB is constitutively expressed on mouse MC. IL-3-dependent mouse bone marrow-derived cultured MC (BMMC) do not express functional FcγRIIIA unless SCF is supplied (8). This has led to the understanding that mouse mucosal MC only express FcγRIIB, whereas connective tissue MC express both FcγRIIB and FcγRIIIA (1). Genetic evidence indicates that in the extracellular portion, mouse FcγRIIIA is most closely related to human FcγRIIA. In conclusion, depending on tissue locations some MC express both the inhibitory receptor (FcγRIIB) and the activating receptor (FcγRIIA for humans and FcγRIIIA for mice), while some MC express one of these.
Both FcγRI and FcγRIIIA require one IgG-binding a-chain and a signal-transducing adaptor composed of a homodimer of the common FcRg-chains for expression and signalling. In contrast, FcγRIIA and FcγRIIB signalling units contain only a single α-chain. The α-chain has three (FcγRI) or two (FcγRIIA, FcγRIIB and FcγRIIIA) immunoglobulin-like extracellular domains that bind the Fc portion of IgG. In addition to an extracellular domain and a transmembrane domain that are shared by all FcγR a-chains, the a-chain of FcγRIIA and FcγRIIB possess, respectively, an intracytoplasmic signaling domain bearing an immunoreceptor tyrosine-based activation motif (ITAM) enabling the coupling to activating protein tyrosine kinases such as spleen tyrosine kinase (Syk) or an immunoreceptor tyrosine-based inhibitory motif (ITIM) enabling the coupling to inhibitoy phosphatases such as src-homology 2-containing inositol 5′ phosphatase (SHIP). The other two MC FcγR, i.e. FcγRI and FcγRIIIA, signal through ITAM located in the FcRγ-chains. In addition to the common g-chain, FcγRIIIA also contains a signal amplifying β-chain that is also shared with FceRI. In contrast to FcRγ widely expressed in hematopoietic cells, FcRβ expression is restricted to MC and basophils. The two extracellular domains of mouse FcγRIIB and FcγRIIIA are 95% identical, and high sequence similarity is also found between human FcγRIIB and FcγRIIA (1).
FcγRI is the only known high-affinity receptor in humans. This receptor binds with a high-affinity only certain subclasses of IgG. For example, it binds IgG2a in mice or IgG1, IgG3 and IgG4 in humans with a high affinity reaching a Ka of about 108 M-1 (9). In contrast, FcγRIIA, FcγRIIB and FcγRIIIA bind IgG with lower affinities (106 to 105 M-1) but with a broader IgG subclass specificity (9). In mice, FcγRIIB and FcγRIIIA can bind all IgG subclasses except IgG3. In contrast, FcγRIIA can bind all the human IgG subtypes (9).
Regulation of mast cell biology by FcγR
Cross-linking of activating FcγR by IgG immune complexes (IgG-IC) launches transmembrane signalling by triggering ITAM tyrosine phosphorylation followed by dynamic protein phosphorylation cascades including SRC family of tyrosine kinase (Lyn), Syk, raft-associated transmembrane adapters (LAT and NTAL) and phosphatidylinositol 3-kinase etc (10). Mouse peritoneal MC degranulate upon treatment with IgG1-IC (11). Crosslinking of FcγRIIIA on connective tissue MC results in degranulation and generation of various lipid mediators (12). In contrast, BMMC cultured in IL-3 resemble mucosal MC, which do not express FcγRIIIA, and they internalize aggregated IgG without degranulation (13). Upregulation of FcγRI by interferon-g on human MC followed by treatment with IgG-IC results in the release of histamine and cytokines (14).
Mice lacking the ITIM-bearing inhibitory FcγRIIB demonstrate increased passive cutaneous anaphylaxis reflecting up-regulated MC activity (1). When coaggregated with receptors bearing ITAM, FcγRIIB negatively regulates ITAM-mediated cell activation. Inhibition of IgE- or IgG-mediated MC activation by FcγRIIB has been firmly established (15-17). It was also demonstrated that aggregation of either FcγRIIB or FcγRIIIA induces apoptosis of mouse BMMC (18). Interestingly, MC with a balanced expression of these two receptors are able to resist apoptosis induced by IgG-IC (18).
Although in vitro experiments undoubtedly support that MC respond to IgG receptor stimulation, information about in vivo contributions of IgG-IC in MC activation is less clear. Recent experiments using new models of genetically modified mice with deficiency in MC and basophils, antibody-mediated cell depletion experiments or experiments with mice specifically depleted of MC FcγR challenge a number of effects previously attributed to MC (9) such as autoimmune arthritis models or experimental allergic encephalomyelitis (EAE) (19-20). Furthermore, while IgG-mediated cutaneous anaphylaxis reactions as initially described in the 1950s undoubtedly show an important MC component, IgG-mediated passive systemic anaphylaxis mediated by FcγRIIIA and PAF release was ascribed to basophils after antibody depletion experiments (9). However, other data show that the strong expression of the inhibitory receptor FcγRIIB versus activating FcγRIIIA in mouse and human basophils rather prevents IgG-mediated activation of these cells (21). Intriguingly, FcγRIV-expressing neutrophils have been found to induce both passive and active systemic anaphylaxis (22).
It should be mentioned that doses of specific antibodies and antigens used to achieve these effects in experimental models are generally high and the relationship to human pathophysiology, where one finds a different distribution of receptors and where usually minute amounts of antigen suffice to induce an anaphylactic shock, remains to be established. IgG-mediated anaphylactic shocks rather seem to be the exception although adverse reactions like serum sickness upon repeated treatment with a heterologous serum or the Arthus reactions after vaccination have been reported very early on (23-24). However, in contrast to the IgE-mediated shock observed by Portier and Richet (25) these reactions develop slowly requiring relatively high doses of antigen.
Conclusion
Mast cell heterogeneity is reflected to a large extent also in FcγR expression. These differences are related to the tissue distribution and maturation state of mast cells. The complexity of the FcγR system and the obvious discrepancies between different species precludes the extrapolation of data obtained in mouse models regarding the effects of FcγR on mast cell biology directly to the human.
References:
| 1. | Malbec, O. and M. Daeron. 2007. The mast cell IgG receptors and their roles in tissue inflammation. Immunol. Rev. 217:206-221. |
| 2. | Bruhns, P. 2012. Properties of mouse and human IgG receptors and their contribution to disease models. Blood. 119:5640-5649. |
| 3. | Smith, K. G. and M. R. Clatworthy. 2010. FcgammaRIIB in autoimmunity and infection: evolutionary and therapeutic implications. Nat Rev Immunol. 10:328-343. |
| 4. | Okayama, Y., A. S. Kirshenbaum, and D. D. Metcalfe. 2000. Expression of a functional high-affinity IgG receptor, Fc gamma RI, on human mast cells: Up-regulation by IFN-gamma. J. Immunol. 164:4332-4339. |
| 5. | Tkaczyk, C., Y. Okayama, M. R. Woolhiser, D. D. Hagaman, A. M. Gilfillan et al. 2002. Activation of human mast cells through the high affinity IgG receptor. Mol. Immunol. 38:1289-1293. |
| 6. | Zhao, W., C. L. Kepley, P. A. Morel et al. 2006. Fc gamma RIIa, not Fc gamma RIIb, is constitutively and functionally expressed on skin-derived human mast cells. J. Immunol. 177:694-701. |
| 7. | Kepley, C. L., S. Taghavi, G. Mackay et al. 2004. Co-aggregation of FcgammaRII with FcepsilonRI on human mast cells inhibits antigen-induced secretion and involves SHIP-Grb2-Dok complexes. J. Biol. Chem. 279:35139-35149. |
| 8. | Katz, H. R. and R. B. Lobell. 1995. Expression and function of Fc gamma R in mouse mast cells. Int Arch Allergy Immunol. 107:76-78. |
| 9. | Jonsson, F. and M. Daeron. 2012. Mast cells and company. Front Immunol. 3:16. |
| 10. | Nimmerjahn, F. and J. V. Ravetch. 2008. Fcgamma receptors as regulators of immune responses. Nat. Rev. Immunol. 8:34-47. |
| 11. | Vaz, N. M. and Z. Ovary. 1968. Passive anaphylaxis in mice with gamma-G antibodies. 3. Release of histamine from mast cells by homologous antibodies. J Immunol. 100:1014-1019. |
| 12. | Katz, H. R., M. B. Raizman, C. S. Gartner, H. C. Scott, A. C. Benson, and K. F. Austen. 1992. Secretory granule mediator release and generation of oxidative metabolites of arachidonic acid via Fc-IgG receptor bridging in mouse mast cells. J Immunol. 148:868-871. |
| 13. | Lobell, R. B., K. F. Austen, and H. R. Katz. 1994. Fc gamma R-mediated endocytosis and expression of cell surface Fc gamma RIIb1 and Fc gamma RIIb2 by mouse bone marrow culture-derived progenitor mast cells. J. Immunol. 152:811-818. |
| 14. | Tkaczyk, C., Y. Okayama, D. D. Metcalfe et al. 2004. Fcgamma receptors on mast cells: activatory and inhibitory regulation of mediator release. Int. Arch. Allergy Immunol. 133:305-315. |
| 15. | Daeron, M., O. Malbec, S. Latour et al. 1995. Regulation of high-affinity IgE receptor-mediated mast cell activation by murine low-affinity IgG receptors. J Clin Invest. 95:577-585. |
| 16. | Takai, T., M. Ono, M. Hikida, H. Ohmori, and J. V. Ravetch. 1996. Augmented humoral and anaphylactic responses in Fc gamma RII-deficient mice. Nature. 379:346-349. |
| 17. | Malbec, O., K. Roget, C. Schiffer et al. 2007. Peritoneal cell-derived mast cells: an in vitro model of mature serosal-type mouse mast cells. J Immunol. 178:6465-6475. |
| 18. | Fang, Y., L. Larsson, P. Bruhns, and Z. Xiang. 2012. Apoptosis of mouse mast cells is reciprocally regulated by the IgG receptors FcgammaRIIB and FcgammaRIIIA. Allergy. 67:1233-1240. |
| 19. | Zhou, J. S., W. Xing, D. S. Friend, K. F. Austen, and H. R. Katz. 2007. Mast cell deficiency in Kit(W-sh) mice does not impair antibody-mediated arthritis. J Exp Med. 204:2797-2802. |
| 20. | Feyerabend, T. B., A. Weiser, A. Tietz et al. 2011. Cre-mediated cell ablation contests mast cell contribution in models of antibody- and T cell-mediated autoimmunity. Immunity. 35:832-844. |
| 21. | Cassard, L., F. Jonsson, S. Arnaud, and M. Daeron. 2012. Fcgamma receptors inhibit mouse and human basophil activation. J Immunol. 189:2995-3006. |
| 22. | Jonsson, F., D. A. Mancardi, Y. Kita et al. 2011. Mouse and human neutrophils induce anaphylaxis. J. Clin. Invest. 121:1484-1496. |
| 23. | Pirquet, C. v. and B. Schick. 1905. Die Serum Krankheit. Serum Sickness (English translation, 1951). Baltimore: Williams and Wilkins Company. |
| 24. | Arthus, M. 1903. Injections répétées de serum de cheval chez le lapin. Compt Rendu Soc de Biol. 50:20. |
| 25. | Portier, P. and C. Richet. 1902. Action anaphylactique des quelques venims. CR Soc Biol. 54:170-172. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}