General description
Mast cells (MC) are best known for their crucial role in the allergic response, but their strategic location and richness of pre-stored and newly synthesized mediators have clearly demonstrated their significance also in innate and adaptive immune responses, inflammation, and tissue remodeling. MC immunomodulatory roles may result in either negative or positive outcome for the host, enhancing, or suppressing certain features of immune/inflammatory responses. MC express a variety of receptors [recently reviewed in (1)] that enable them, both in activating or inhibiting ways, to modulate immune responses and interact with other cells and molecules, and can be used as treatment targets in disease conditions. The activating receptors are comprised of many types, differentiated by their ligands, morphology and function such as chemokines (2), interleukins (IL)(3), amines (4), Toll-Like Receptors (TLRs)(5), purinergic (6) and others (see references for examples). The activating receptors work via different mechanisms, result in one or more of the following: degranulation, migration, production and release of cytokines, adhesion etc.
MC function can also be greatly affected by cross-talk between neighboring receptors on the surface of the same MC or between receptors on diverse cell types, which might result in co-activation (synergism or additive effects). A few examples of activating co-stimulation between FcεRI, the main activating receptor of MC which is also used as a fingerprint in those cells, and adenosine receptor or the chemokine receptor CCRI, were recently reviewed in (1). Co-stimulation was shown to be often required for the development of an effective immune response, and has been recognized to participate in antigen-specific signal in lymphocytes antigen receptors (e.g (1, 7.8)).
CD48 as a representative example
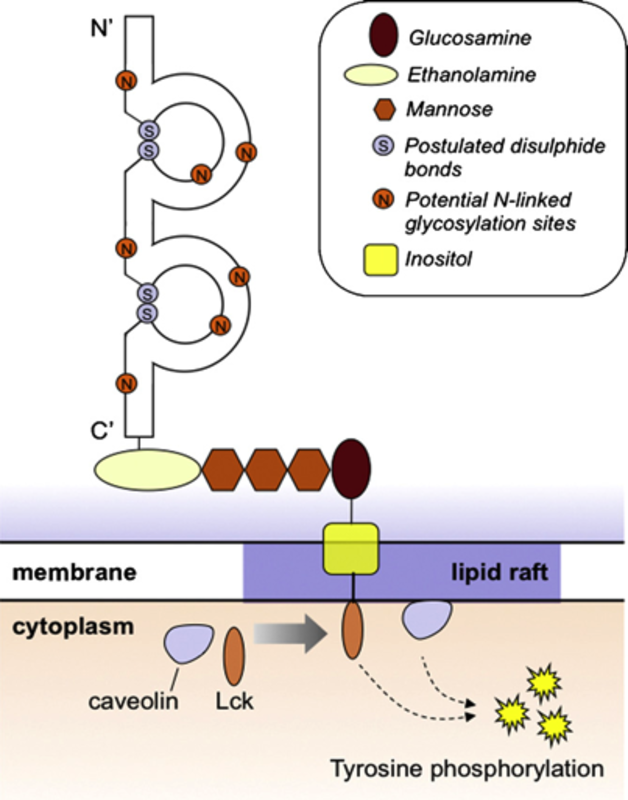
Characteristics: CD48, aCD2-like molecule, is a 40KD glycosyl-phosphatidyl- inositol (GPI) anchored protein, expressed on the surface of hematopoietic cells (recently reviewed in (9)). The CD48 molecule has a distal V-like domain with a C2-like domain containing conserved cysteine residues that form disulfide bonds. It lacks a trans membrane domain and is attached to the cell surface by a glycolipid, GPI, restricted to the outer leaflet of the membrane bilayer (10). CD48 has also a soluble form found upon activation due to its cleavage. Stimulated CD48 associates to the kinase LCK and leads to tyrosine phosphorylation (9).
Expression and function: In MC, CD48 was first described to be involved in innate immunity as it binds FimH of E. coli. This interaction triggers a strong TNFα release and the uptake of this Gram- bacteria (11, 12, 13). It was also shown to participate in MC interactions with M. tuberculosis, which triggers the release of several pre-stored mediators and the synthesis of different cytokines (14). S. aureus, a Gram+ bacteria, and its toxins were later shown to bind CD48 and TLR2 on human cord blood MC (hCBMC) causing subsequent release of pro-inflammatory cytokines (15).
The physical interactions of MC CD48 with its human high affinity ligand 2B4 on eosinophils, play an important role in allergic inflammation due to the formation of the allergic effector unit (AEU) between the two cells (16). CD48-2B4 binding induces degranulation of MC and increases eosinophil survival and activation (16). CD48 is over expressed in the murine asthma model and was defined as a signature gene in this condition (17).
Pharmacology
Evidences show that MC receptors have a strong therapeutic potential but still a better understanding of their pathophysiological roles, expression and their role in the immune system is required. As an important activating MC receptor, both by itself and as part of the AEU, CD48 is likely to be further studied for its use as a therapeutic target, in order to avoid both AEU formation and bacterial invasion. In a murine model of allergic asthma, treatment with neutralizing CD48 Ab dramatically inhibited the lung inflammation (18).
References:
| 1. | Migalovich-Sheikhet H, Friedman S, Mankuta D, Levi-Schaffer F. 2012. Novel identified receptors on MC. Front in Immunol 3:238. |
| 2. | Lacotte S, Brun S, Muller S, Dumortier H. 2009. CXCR3, inflammation, and autoimmune diseases. Ann N Y Acad Sci. 1173:310-7. |
| 3. | Akdis M, Burgler S, Crameri R, Eiwegger T, Fujita H, Gomez E, Klunker S, Meyer N, O’Mahony L, PalomaresO, Rhyner C, Ouaked N, Schaffartzik A, vandeVeen W, Zeller S, Zimmermann M, and Akdis C.A. 2011. Interleukins, from1to37, andinterferon-gamma: receptors, functions, and roles in diseases. J. AllergyClin.Immunol. 127:701–21. |
| 4. | Gibbs BF and Levi-Schaffer F. 2012. H₄ receptors in mast cells and basophils: a new therapeutic target for allergy? Front Bio Sci. 17:430-7. |
| 5. | Supajatura V, Ushio H, Nakao A, Akira S, Okumura K, Ra C, Ogawa H. 2002. Differential responses of mast cell Toll-like receptors 2 and 4 in allergy and innate immunity. J Clin Invest. 109(10):1351-9. |
| 6. | Bulanova E, and Bulfone-Paus S. 2010. P2 receptor-mediated signaling in mast cell biology. Purinergic Signal. 6(1):3-17. |
| 7. | Nunomura S, Gon Y, Yoshimaru T, Kashiwakura J, Kawakami T, Ra C. 2010. FcepsilonRI beta-chain ITAM amplifies PI3K-signaling to ensure synergistic degranulation response via FcepsilonRI and adenosine receptors. Eur J Immunol. 40(4):1205-17. |
| 8. | Aye CC, Toda M, Morohoshi K, Ono SJ. 2012. Identification of genes and proteins specifically regulated by costimulation of mast cell Fcε Receptor I and chemokine receptor 1. Exp Mol Pathol. 92(3):267-74. |
| 9. | Elishmereni M and Levi- Schaffer F. 2011. CD48: A co-stimulatory receptor of immunity. Int J Biochem Cell Biol. 43(1):25-8. |
| 10. | Shin JS, Abraham SN. 2001. Glycosylphosphatidylinositol-anchored receptor-mediated bacterial endocytosis. FEMS Microbiol Lett. 13;197(2):131-8. |
| 11. | Malaviya R, Gao Z, Thankavel K, van der Merwe PA, Abraham SN. 1999. The mast cell tumor necrosis factor alpha response to FimH-expressing Escherichia coli is mediated by the glycosylphosphatidylinositol-anchored molecule CD48. Proc Natl Acad Sci U S A. 96(14):8110-5. |
| 12. | Malaviya R, Abraham SN.2001. Mast cell modulation of immune responses to bacteria. Immunol Rev. 179:16-24. |
| 13. | Proft T, Baker EN. 2009. Pili in Gram-negative and Gram-positive bacteria – structure, assembly and their role in disease. Cell Mol Life Sci. 66(4):613-35. |
| 14. | Muñoz S, Hernández-Pando R, Abraham SN, Enciso JA.2003. Mast cell activation by Mycobacterium tuberculosis: mediator release and role of CD48. J Immunol. 170(11):5590-6. |
| 15. | Rocha-de-Souza CM, Berent-Maoz B, Mankuta D, Moses AE, Levi-Schaffer F. 2008. Human mast cell activation by Staphylococcus aureus: interleukin-8 and tumor necrosis factor alpha release and the role of Toll-like receptor 2 and CD48 molecules. Infect Immun. 76(10):4489-97. |
| 16. | Elishmereni M, Alenius HT, Bradding P, Mizrahi S, Shikotra A, Minai-Fleminger Y, Mankuta D, Eliashar R, Zabucchi G, Levi-Schaffer F. 2011. Physical interactions between mast cells and eosinophils: a novel mechanism enhancing eosinophil survival in vitro. Allergy. 66(3):376-85. |
| 17. | Zimmermann N, King NE, Laporte J, Yang M, Mishra A, Pope SM, Muntel EE, Witte DP, Pegg AA, Foster PS, Hamid Q, Rothenberg ME. 2003. Dissection of experimental asthma with DNA microarray analysis identifies arginase in asthma pathogenesis. J Clin Invest. 111(12):1863-74. |
| 18. | Munitz A, Bachelet I, Finkelman FD, Rothenberg ME, Levi-Schaffer F. 2007. CD48 is critically involved in allergic eosinophilic airway inflammation. Am J Respir Crit Care Med. 175(9):911-8. |


{kind=link}