General description
Basophils are relatively rare cells that make up less than 1 % of blood leukocytes and are thought to play a role in expelling certain parasites but also contribute to allergic diseases. They were discovered in 1891 by Paul Ehrlich who showed that these granulated cells display similar staining characteristics to mast cells. Like mast cells, basophils produce histamine, which is stored in granules [1] consisting of proteoglycans from which it is released following stimulation and causes inflammation. Although basophils share many other properties with mast cells, such as expressing high-affinity IgE receptors (FcεRI), evidence to date suggests these cells are derived from different myeloid progenitors in the bone marrow (which also lead to eosinophils) [2] and certain key functional differences though these vary depending on the species. Basophils are generated in the bone marrow from where they are released into the blood. Recent advances in immunostaining (e.g. using antibodies to basogranulin, a basophil-specific protein) [3] has now demonstrated that basophils invade other tissues, such as the lungs and skin, or migrate to lymph nodes, during allergic inflammation and parasitic infections [4,5].
Activation of basophils
Basophils can be activated by nanograms per liter concentrations of allergen following prior sensitization with allergen-specific IgE. Activation occurs by crosslinking of at least two (probably more) IgE-FcεRI complexes with allergen, triggering the phosphorylation of Lyn and Syk protein kinases which control subsequent activation of stimulatory signal transduction pathways which crucially involve the influx of calcium ions into basophils. Lyn is also associated with the initiation of inhibitory signaling pathways involving phosphatases like SHIP-1, which limits basophil activation [6].
There are a number of other stimuli that cause non-specific crosslinking of FcεRI-bound IgE, including the IPSE/alpha-1 present in Schistosoma mansoni eggs [7], HIV-1 gp120 [8], B cell superantigens [9], leading to the introduction of the term ‘superallergen’ [10], and dietary lectins [11]. Stimulation of such IgE-dependent stimuli can be markedly enhanced by a number of priming cytokines, such as IL-3 [12], GM-CSF [13], NGF [14] and IL-33 [15], as well as chemokines such as eotaxin [16]. IgE-independent activators of basophils include the gram positive bacterial peptide fMLP [17] as well as complement factors C5a [18] and C3a [19], which can also cause marked degranulation, TLR ligands (although these do not usually cause substantial histamine release) [20], and leukocyte immunoglobulin-like receptor (LIR)-7 [21]. Basophil activation also results in the fast up-regulation of several surface activation markers (Table 1). These can be used for detection of basophil activation in basophil activation tests (BATs) in whole blood [22–24].
Basophil-derived mediators
Degranulation of basophils results in the rapid release of histamine which usually occurs within 30 minutes of allergen stimulation (and more rapidly for fMLP). This is followed immediately by the production and release of the eicosanoid leukotriene C4 (but unlike mast cells basophils do not produce cyclooxygenase products such as prostaglandin D2) [25]. Interestingly, basophils rapidly release IL-4 within 4 hours after IgE-dependent stimulation and IL-13 over longer periods [26,27]. Both cytokines play a central role in supporting late-phase allergic reactions (by upregulating vascular adhesion molecules) and pro-allergic Th2 immunity. It is thought that basophils are the main early source of these cytokines whereas human mast cells are poor producers of these factors, especially IL-4. They have also been reported to release VEGF [28], a factor involved in angiogenesis and tissue remodelling.
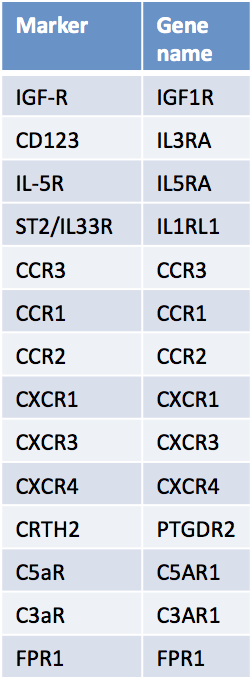
Role of basophils in allergy and parasite immunity
Although basophils are usually found in the circulation and are rarely present in normal tissues, they have been widely shown to invade allergic inflamed sites in allergic individuals. Basophils express various chemokine receptors (see Table 2 for a selection) and respond to many chemotactic agents which aid their migration. Activation of basophils in allergic inflammatory sites has been demonstrated by differential mediator analysis, indicating that basophils and not mast cells do release their mediators in the late phase of the allergic response. Their presence in tissues affected by allergic disease, including in the airways of asthmatic patients, in the skin of atopic dermatitis patients, and in the nose has been demonstrated using basophil-specific BB1 and 2D7 antibodies [3,29]. In the rapid anaphylactic reactions following insect venom exposure, or food intake in food allergic patients, blood basophils and not so much tissue-resident mast cells have been hypothesized to play a role.
It is unlikely that basophils, with their clear potential to have detrimental effects, would have evolved and been widely preserved in higher animals without an important immune function. The first indications of such a role of basophils goes back to the pioneering work of William Trager, who demonstrated an essential involvement of basophils in immunity to hard ticks [30]. A more refined understanding regarding the interplay between basophils and mast cells in tick infestation comes from the work of Hajime Karasuyama and his team [31]. The role of basophil in immunity is not limited to exoparasites. A role for basophils in immunity to endoparasites (intestinal helminths) has been described by many authors since the pioneering work of Jarrett and Bazin [32]. More recently, the availability of murine basophil knockout models has enabled researchers to dissect the roles of basophils in immunity to helminth parasites in greater detail [31,33]. It remains to be determined to which extent these findings can be extrapolated to human basophils [34–36].
References:
| 1. | Sampson D, Archer GT (1967) Release of histamine from human basophils. Blood 29: 722–736. Available: www.ncbi.nlm.nih.gov/pubmed/4164650. |
| 2. | Görgens A, Radtke S, Möllmann M, Cross M, Dürig J, et al. (2013) Revision of the human hematopoietic tree: granulocyte subtypes derive from distinct hematopoietic lineages. Cell Rep 3: 1539–1552. Available: www.ncbi.nlm.nih.gov/pubmed/23707063. |
| 3. | McEuen AR, Buckley MG, Compton SJ, Walls AF (1999) Development and characterization of a monoclonal antibody specific for human basophils and the identification of a unique secretory product of basophil activation. Lab Invest 79: 27–38. Available: www.ncbi.nlm.nih.gov/pubmed/9952108. |
| 4. | Charlesworth EN, Hood AF, Soter NA, Kagey-Sobotka A, Norman PS, et al. (1989) Cutaneous late-phase response to allergen. Mediator release and inflammatory cell infiltration. J Clin Invest 83: 1519–1526. Available: www.pubmedcentral.nih.gov/articlerender.fcgi. |
| 5. | Bascom R, Wachs M, Naclerio RM, Pipkorn U, Galli SJ, et al. (1988) Basophil influx occurs after nasal antigen challenge: effects of topical corticosteroid pretreatment. J Allergy Clin Immunol 81: 580–589. Available: www.ncbi.nlm.nih.gov/pubmed/2450113. |
| 6. | Gibbs BF, Räthling A, Zillikens D, Huber M, Haas H (2006) Initial Fc epsilon RI-mediated signal strength plays a key role in regulating basophil signaling and deactivation. J Allergy Clin Immunol 118: 1060–1067. Available: www.ncbi.nlm.nih.gov/pubmed/17088130. |
| 7. | Schramm G, Falcone FH, Gronow A, Haisch K, Mamat U, et al. (2003) Molecular characterization of an interleukin-4-inducing factor from Schistosoma mansoni eggs. J Biol Chem 278: 18384–18392. Available: www.ncbi.nlm.nih.gov/pubmed/12624091. |
| 8. | Patella V, Florio G, Petraroli A, Marone G (2000) HIV-1 gp120 induces IL-4 and IL-13 release from human Fc epsilon RI+ cells through interaction with the VH3 region of IgE. J Immunol 164: 589–595. Available: www.ncbi.nlm.nih.gov/pubmed/10623799. |
| 9. | Genovese A, Borgia G, Björck L, Petraroli A, de Paulis A, et al. (2003) Immunoglobulin superantigen protein L induces IL-4 and IL-13 secretion from human Fc epsilon RI+ cells through interaction with the kappa light chains of IgE. J Immunol 170: 1854–1861. Available: www.ncbi.nlm.nih.gov/pubmed/12574351. |
| 10. | Marone G, Rossi FW, Detoraki A, Granata F, Marone G, et al. (2007) Role of superallergens in allergic disorders. Chem Immunol Allergy 93: 195–213. Available: www.ncbi.nlm.nih.gov/pubmed/17369707. |
| 11. | Haas H, Falcone FH, Schramm G, Haisch K, Gibbs BF, et al. (1999) Dietary lectins can induce in vitro release of IL-4 and IL-13 from human basophils. Eur J Immunol 29: 918–927. Available: www.ncbi.nlm.nih.gov/pubmed/10092096. |
| 12. | Mayer P, Valent P, Schmidt G, Liehl E, Bettelheim P (1989) The in vivo effects of recombinant human interleukin-3: demonstration of basophil differentiation factor, histamine-producing activity, and priming of GM-CSF-responsive progenitors in nonhuman primates. Blood 74: 613–621. Available: www.ncbi.nlm.nih.gov/pubmed/2665851. |
| 13. | Sarmiento EU, Espiritu BR, Gleich GJ, Thomas LL (1995) IL-3, IL-5, and granulocyte-macrophage colony-stimulating factor potentiate basophil mediator release stimulated by eosinophil granule major basic protein. J Immunol 155: 2211–2221. Available: www.ncbi.nlm.nih.gov/pubmed/7543541. |
| 14. | Bischoff SC, Dahinden CA (1992) Effect of nerve growth factor on the release of inflammatory mediators by mature human basophils. Blood 79: 2662–2669. Available: www.ncbi.nlm.nih.gov/pubmed/1586715. |
| 15. | Pecaric-Petkovic T, Didichenko SA, Kaempfer S, Spiegl N, Dahinden CA (2009) Human basophils and eosinophils are the direct target leukocytes of the novel IL-1 family member IL-33. Blood 113: 1526–1534. Available: www.pubmedcentral.nih.gov/articlerender.fcgi. |
| 16. | Devouassoux G, Metcalfe DD, Prussin C (1999) Eotaxin potentiates antigen-dependent basophil IL-4 production. J Immunol 163: 2877–2882. Available: www.ncbi.nlm.nih.gov/pubmed/10453034. |
| 17. | Conroy MC, Kennedy JA, Thueson DO (1985) Differential inhibition of histamine release from human basophils induced by antigen, anti-IgE or N-formyl-L-methionyl-L-leucyl-L-phenylalanine. Int Arch Allergy Appl Immunol 77: 222–224. Available: www.ncbi.nlm.nih.gov/pubmed/2409021. |
| 18. | Kurimoto Y, de Weck AL, Dahinden CA (1989) Interleukin 3-dependent mediator release in basophils triggered by C5a. J Exp Med 170: 467–479. Available: www.pubmedcentral.nih.gov/articlerender.fcgi. |
| 19. | Bischoff SC, de Weck AL, Dahinden CA (1990) Interleukin 3 and granulocyte/macrophage-colony-stimulating factor render human basophils responsive to low concentrations of complement component C3a. Proc Natl Acad Sci U S A 87: 6813–6817. Available: www.pubmedcentral.nih.gov/articlerender.fcgi. |



{kind=link}
{kind=link}
{kind=link}